

Adaptations for economical bipedal running: the effect of limb structure on three-dimensional joint mechanics
- PMID: 21030429
- PMCID: PMC3061092
- DOI: 10.1098/rsif.2010.0466
Adaptations for economical bipedal running: the effect of limb structure on three-dimensional joint mechanics
Abstract
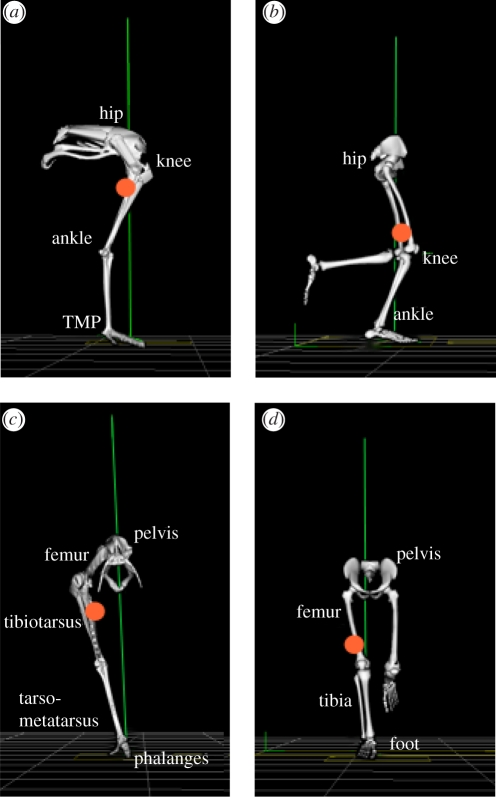
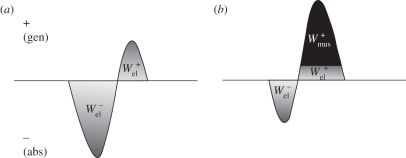
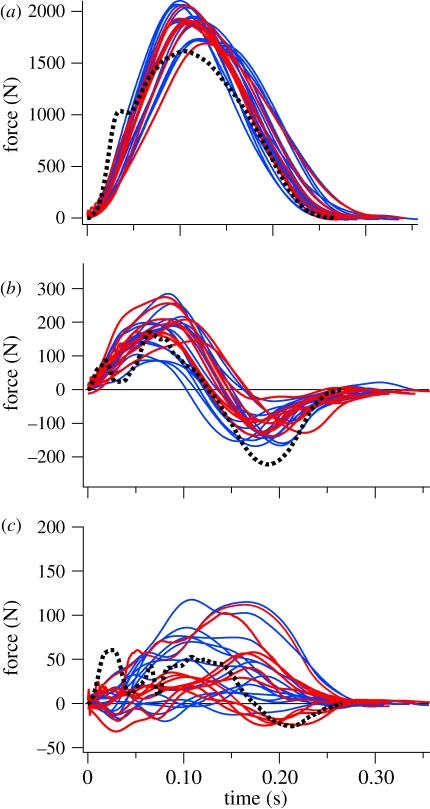
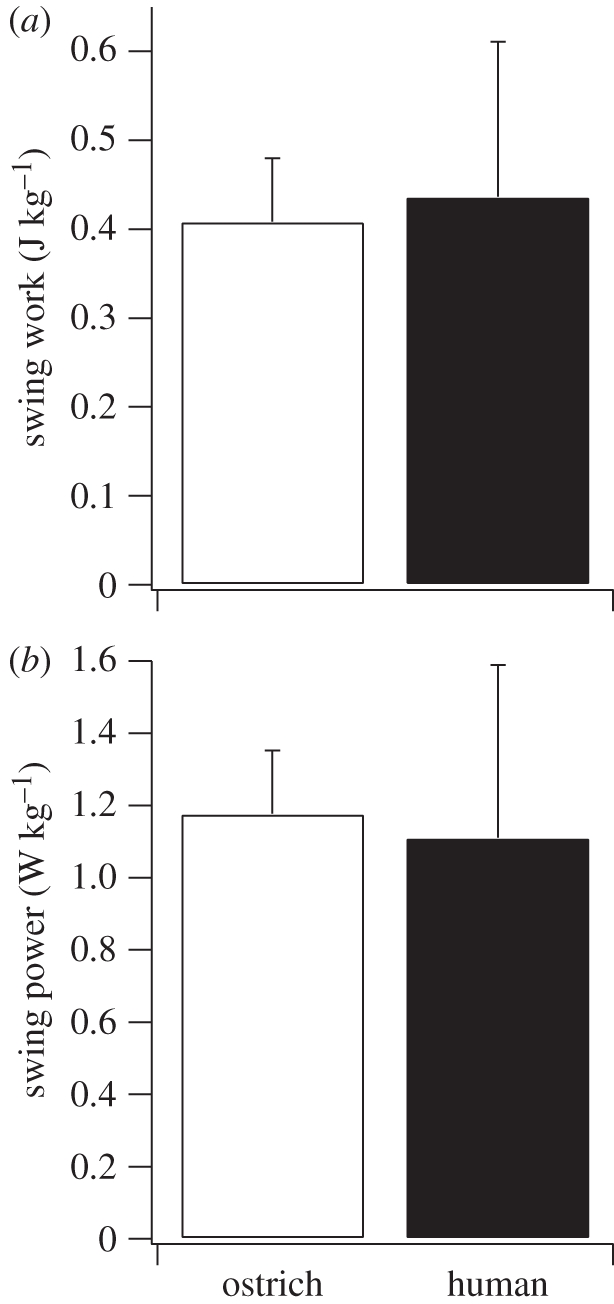
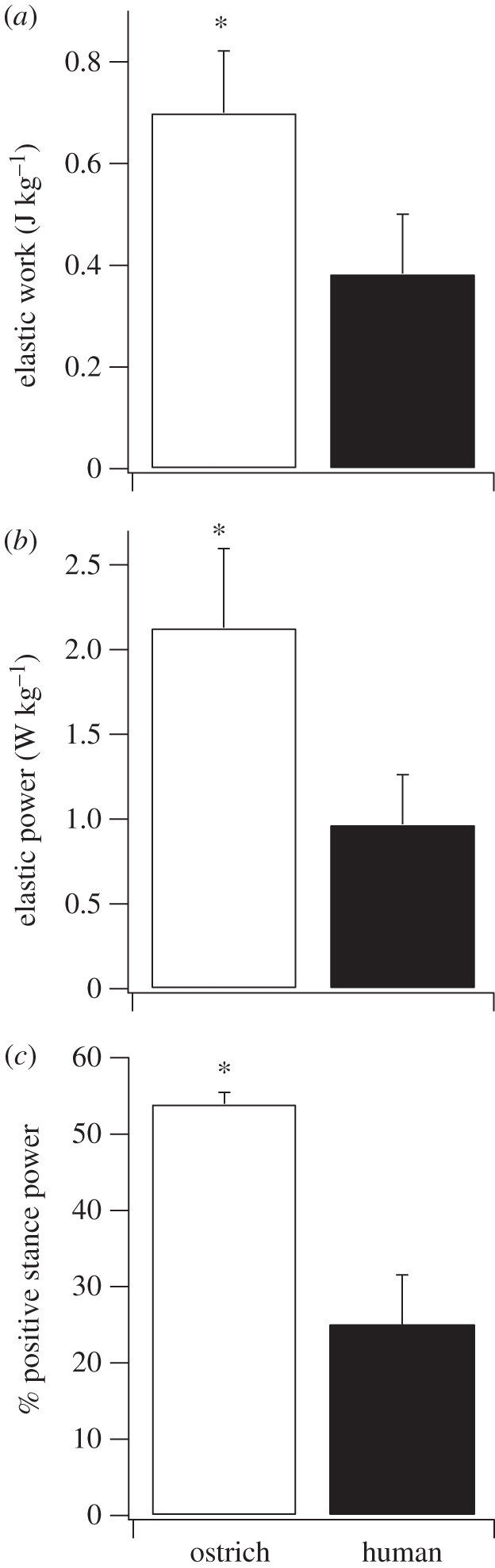
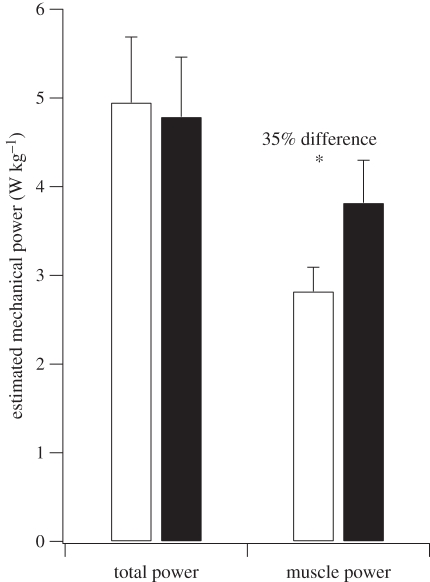
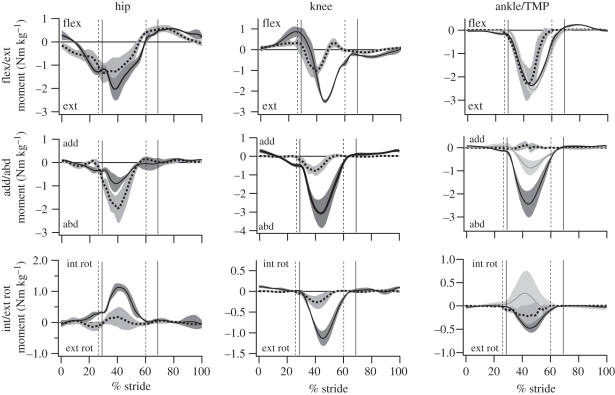
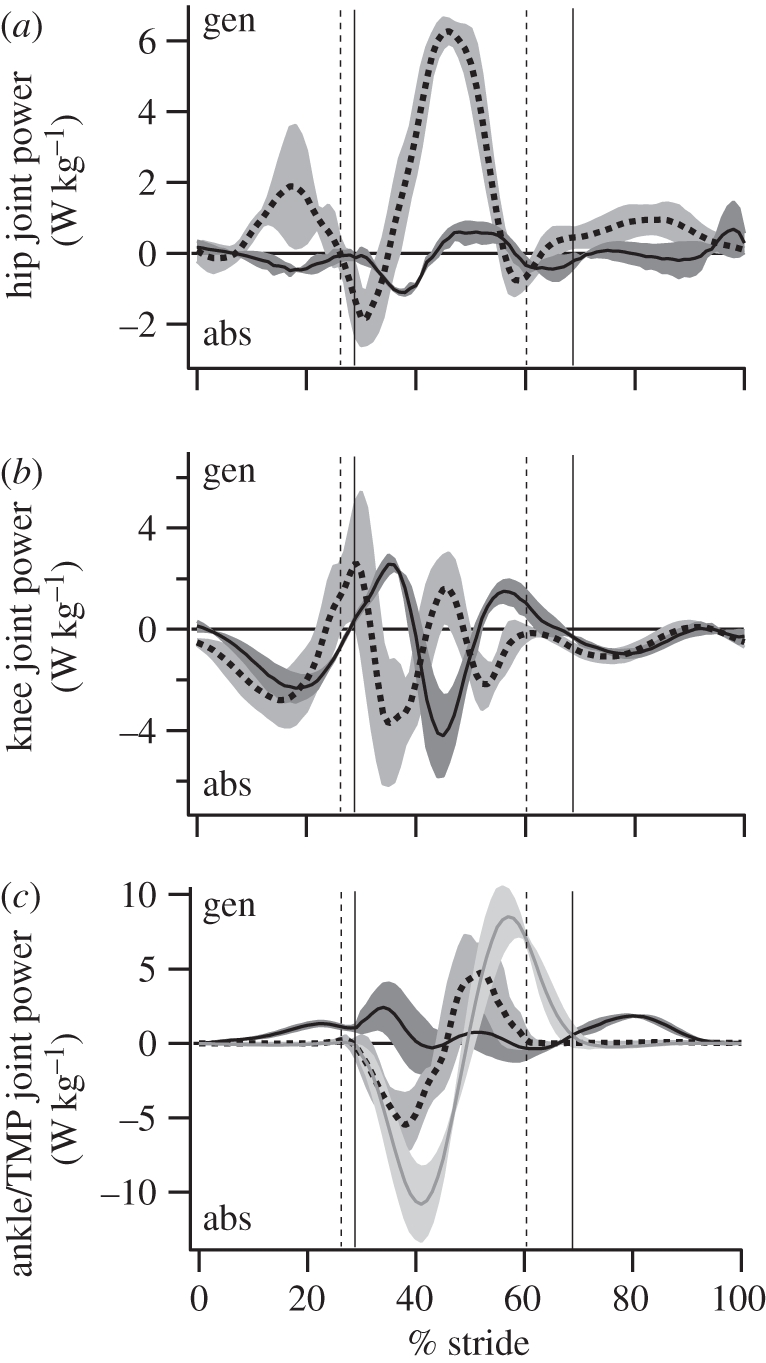
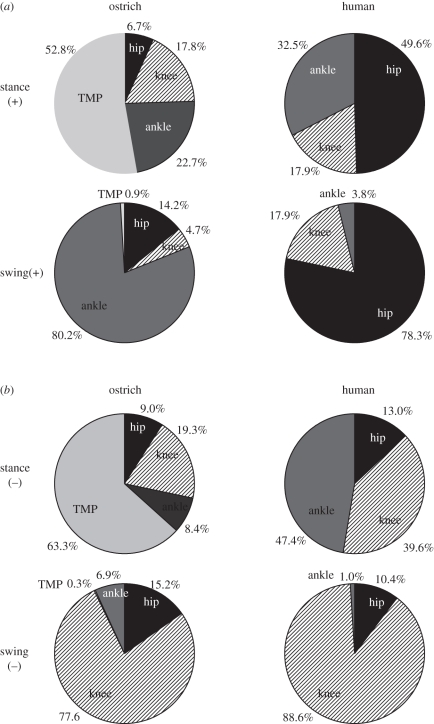
The purpose of this study was to examine the mechanical adaptations linked to economical locomotion in cursorial bipeds. We addressed this question by comparing mass-matched humans and avian bipeds (ostriches), which exhibit marked differences in limb structure and running economy. We hypothesized that the nearly 50 per cent lower energy cost of running in ostriches is a result of: (i) lower limb-swing mechanical power, (ii) greater stance-phase storage and release of elastic energy, and (iii) lower total muscle power output. To test these hypotheses, we used three-dimensional joint mechanical measurements and a simple model to estimate the elastic and muscle contributions to joint work and power. Contradictory to our first hypothesis, we found that ostriches and humans generate the same amounts of mechanical power to swing the limbs at a similar self-selected running speed, indicating that limb swing probably does not contribute to the difference in energy cost of running between these species. In contrast, we estimated that ostriches generate 120 per cent more stance-phase mechanical joint power via release of elastic energy compared with humans. This elastic mechanical power occurs nearly exclusively at the tarsometatarso-phalangeal joint, demonstrating a shift of mechanical power generation to distal joints compared with humans. We also estimated that positive muscle fibre power is 35 per cent lower in ostriches compared with humans, and is accounted for primarily by higher capacity for storage and release of elastic energy. Furthermore, our analysis revealed much larger frontal and internal/external rotation joint loads during ostrich running than in humans. Together, these findings support the hypothesis that a primary limb structure specialization linked to economical running in cursorial species is an elevated storage and release of elastic energy in tendon. In the ostrich, energy-saving specializations may also include passive frontal and internal/external rotation load-bearing mechanisms.
Figures
Similar articles
-
Adaptations for bipedal walking: Musculoskeletal structure and three-dimensional joint mechanics of humans and bipedal chimpanzees (Pan troglodytes).J Hum Evol. 2022 Jul;168:103195. doi: 10.1016/j.jhevol.2022.103195. Epub 2022 May 18. J Hum Evol. 2022. PMID: 35596976
-
Mechanical and energetic scaling relationships of running gait through ontogeny in the ostrich (Struthio camelus).J Exp Biol. 2013 Mar 1;216(Pt 5):841-9. doi: 10.1242/jeb.064691. Epub 2012 Nov 15. J Exp Biol. 2013. PMID: 23155079
-
Inferring muscle functional roles of the ostrich pelvic limb during walking and running using computer optimization.J R Soc Interface. 2016 May;13(118):20160035. doi: 10.1098/rsif.2016.0035. J R Soc Interface. 2016. PMID: 27146688 Free PMC article.
-
Patterns of mechanical energy change in tetrapod gait: pendula, springs and work.J Exp Zool A Comp Exp Biol. 2006 Nov 1;305(11):899-911. doi: 10.1002/jez.a.334. J Exp Zool A Comp Exp Biol. 2006. PMID: 17029267 Review.
-
Contributions to the understanding of gait control.Dan Med J. 2014 Apr;61(4):B4823. Dan Med J. 2014. PMID: 24814597 Review.
Cited by
-
Hindlimb kinematics, kinetics and muscle dynamics during sit-to-stand and sit-to-walk transitions in emus (Dromaius novaehollandiae).J Exp Biol. 2024 Dec 15;227(24):jeb247519. doi: 10.1242/jeb.247519. Epub 2024 Dec 17. J Exp Biol. 2024. PMID: 39445465 Free PMC article.
-
Computational modelling of muscle fibre operating ranges in the hindlimb of a small ground bird (Eudromia elegans), with implications for modelling locomotion in extinct species.PLoS Comput Biol. 2021 Apr 1;17(4):e1008843. doi: 10.1371/journal.pcbi.1008843. eCollection 2021 Apr. PLoS Comput Biol. 2021. PMID: 33793558 Free PMC article.
-
The Bionic High-Cushioning Midsole of Shoes Inspired by Functional Characteristics of Ostrich Foot.Bioengineering (Basel). 2022 Dec 20;10(1):1. doi: 10.3390/bioengineering10010001. Bioengineering (Basel). 2022. PMID: 36671573 Free PMC article.
-
Swing-leg trajectory of running guinea fowl suggests task-level priority of force regulation rather than disturbance rejection.PLoS One. 2014 Jun 30;9(6):e100399. doi: 10.1371/journal.pone.0100399. eCollection 2014. PLoS One. 2014. PMID: 24979750 Free PMC article.
-
Elite long jumpers with below the knee prostheses approach the board slower, but take-off more effectively than non-amputee athletes.Sci Rep. 2017 Nov 22;7(1):16058. doi: 10.1038/s41598-017-16383-5. Sci Rep. 2017. PMID: 29167568 Free PMC article.
References
-
- Carrano M. T. 1999. What, if anything, is a cursor? Categories versus continua for determining locomotor habit in mammals and dinosaurs. J. Zool. Lond. 247, 29–4210.1111/j.1469-7998.1999.tb00190.x (doi:10.1111/j.1469-7998.1999.tb00190.x) - DOI - DOI
-
- Rubenson J., Heliams D. B., Maloney S. K., Withers P. C., Lloyd D. G., Fournier P. A. 2007. Reappraisal of the comparative cost of human locomotion using gait-specific allometric analyses. J. Exp. Biol. 210, 3513–352410.1242/jeb.000992 (doi:10.1242/jeb.000992) - DOI - DOI - PubMed
-
- Biewener A. A. 1998. Muscle function in vivo: a comparison of muscles used for elastic energy savings versus muscles used to generate mechanical power. Am. Zool. 38, 703–717
-
- Steudel K. 1990. The work and energetic cost of locomotion. II. Partitioning the cost of internal and external work within a species. J. Exp. Biol. 154, 287–303 - PubMed
-
- Wilson A. M., Van den Bogert A. J., Mcguigan M. P. 2000. Optimization of the muscle–tendon unit for economical locomotion in cursorial animals. In Skeletal muscle mechanics: from mechanism to function (ed. Herzog W.), pp. 517–547 Hoboken, NJ: John Wiley & Sons
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
