

Reverse-genetic analysis of the two biotin-containing subunit genes of the heteromeric acetyl-coenzyme A carboxylase in Arabidopsis indicates a unidirectional functional redundancy
- PMID: 21030508
- PMCID: PMC3075786
- DOI: 10.1104/pp.110.165910
Reverse-genetic analysis of the two biotin-containing subunit genes of the heteromeric acetyl-coenzyme A carboxylase in Arabidopsis indicates a unidirectional functional redundancy
Abstract
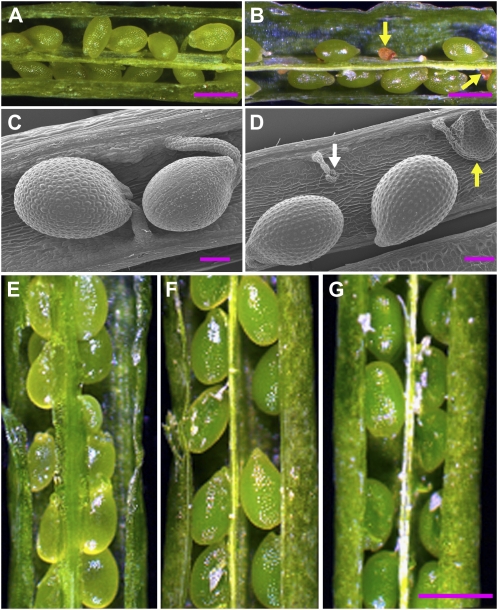
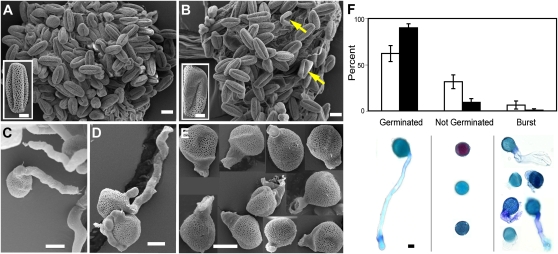
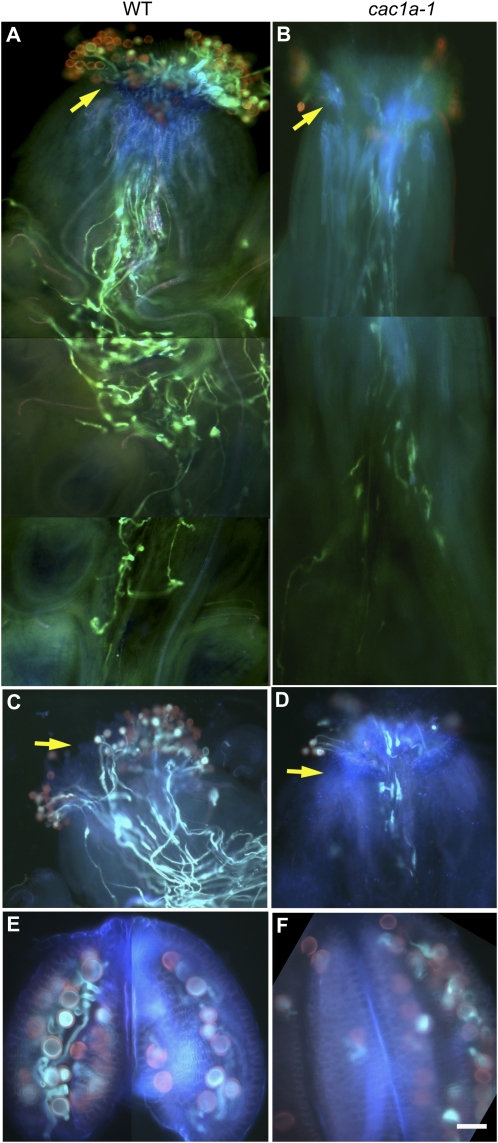
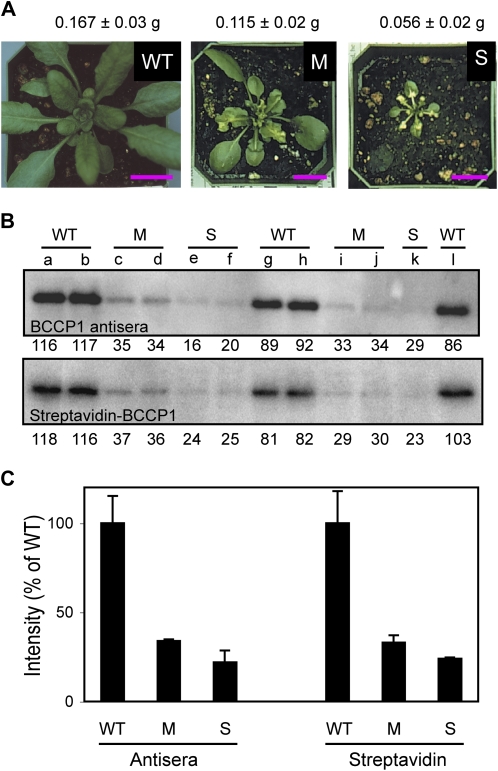
The heteromeric acetyl-coenzyme A carboxylase catalyzes the first and committed reaction of de novo fatty acid biosynthesis in plastids. This enzyme is composed of four subunits: biotin carboxyl-carrier protein (BCCP), biotin carboxylase, α-carboxyltransferase, and β-carboxyltransferase. With the exception of BCCP, single-copy genes encode these subunits in Arabidopsis (Arabidopsis thaliana). Reverse-genetic approaches were used to individually investigate the physiological significance of the two paralogous BCCP-coding genes, CAC1A (At5g16390, codes for BCCP1) and CAC1B (At5g15530, codes for BCCP2). Transfer DNA insertional alleles that completely eliminate the accumulation of BCCP2 have no perceptible effect on plant growth, development, and fatty acid accumulation. In contrast, transfer DNA insertional null allele of the CAC1A gene is embryo lethal and deleteriously affects pollen development and germination. During seed development the effect of the cac1a null allele first becomes apparent at 3-d after flowering, when the synchronous development of the endosperm and embryo is disrupted. Characterization of CAC1A antisense plants showed that reducing BCCP1 accumulation to 35% of wild-type levels, decreases fatty acid accumulation and severely affects normal vegetative plant growth. Detailed expression analysis by a suite of approaches including in situ RNA hybridization, promoter:reporter transgene expression, and quantitative western blotting reveal that the expression of CAC1B is limited to a subset of the CAC1A-expressing tissues, and CAC1B expression levels are only about one-fifth of CAC1A expression levels. Therefore, a likely explanation for the observed unidirectional redundancy between these two paralogous genes is that whereas the BCCP1 protein can compensate for the lack of BCCP2, the absence of BCCP1 cannot be tolerated as BCCP2 levels are not sufficient to support heteromeric acetyl-coenzyme A carboxylase activity at a level that is required for normal growth and development.
Figures








References
-
- Alexander MP. (1969) Differential staining of aborted and nonaborted pollen. Stain Technol 44: 117–122 - PubMed
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Bao X, Shorrosh BS, Ohlrogge JB. (1997) Isolation and characterization of an Arabidopsis biotin carboxylase gene and its promoter. Plant Mol Biol 35: 539–550 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
