

Review
doi: 10.1016/B978-0-12-381300-8.00002-2.
Innate signaling networks in mucosal IgA class switching
Affiliations
- PMID: 21034970
- PMCID: PMC3046556
- DOI: 10.1016/B978-0-12-381300-8.00002-2
Item in Clipboard
Review
Innate signaling networks in mucosal IgA class switching
Adv Immunol.
2010.
Abstract
The past 20 years have seen a growing interest over the control of adaptive immune responses by the innate immune system. In particular, considerable attention has been paid to the mechanisms by which antigen-primed dendritic cells orchestrate the differentiation of T cells. Additional studies have elucidated the pathways followed by T cells to initiate immunoglobulin responses in B cells. In this review, we discuss recent advances on the mechanisms by which intestinal bacteria, epithelial cells, dendritic cells, and macrophages cross talk with intestinal T cells and B cells to induce frontline immunoglobulin A class switching and production.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
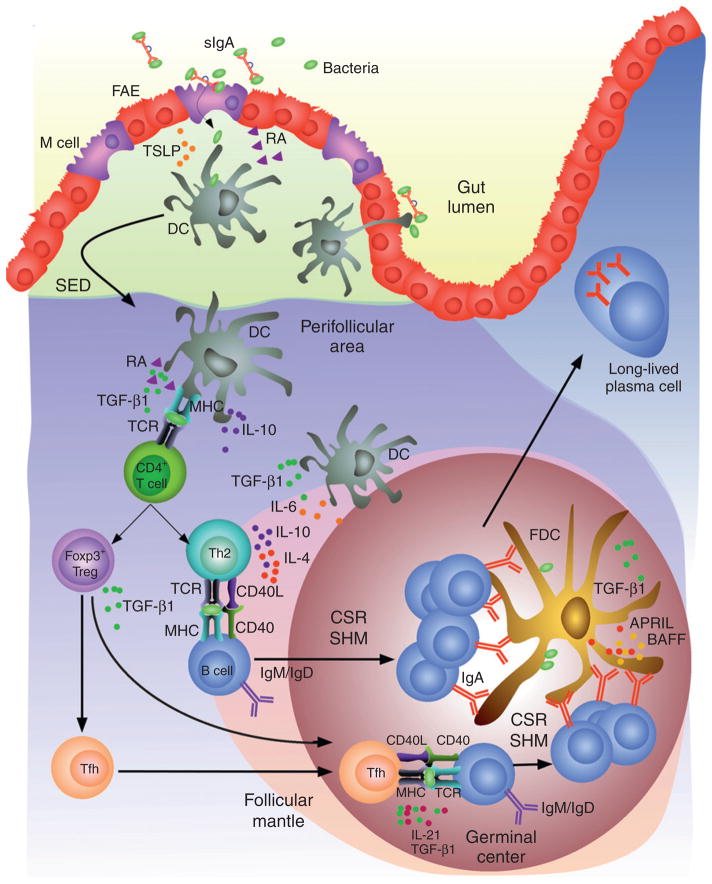
TD induction of IgA CSR and production in intestinal Peyer’s patches. DCs located in the subepithelial dome (SED) of Peyer’s patches capture SIgA-bound or free microbial antigens by interacting with M cells or by extending dendritic projections into the lumen of the gut. While capturing antigen, DCs are exposed to “conditioning” factors such as TSLP and RA, which are produced by IECs in response to microbial TLR ligands. TSLP and RA prevent antigen-loaded DCs from inducing inflammatory Th1 responses. Instead, TSLP- and RA-primed DCs migrate to the perifollicular area to promote differentiation of naïve CD4+ T cells into Treg and Th2 cells. These noninflammatory T cells induce IgA CSR and production by activating B cells through CD40L and cytokines such as IL-4, IL-10, and TGF-β in the context of an antigen-driven cognate T–B cell interaction. TSLP- and RA-primed DCs further enhance IgA CSR and production by releasing IL-6, IL-10, and TGF-β1. Of note, Treg cells can further differentiate into Tfh cells, which trigger IgA CSR and production through IL-21 and TGF-β1. In the germinal center of Peyer’s patches, B cells undergo SHM in addition to completing IgA CSR. In the presence of antigen-exposing FDCs, SHM facilitates the selection of B cells expressing high-affinity IgA. FDCs also enhance IgA CSR and production by releasing BAFF, APRIL, and TGF-β1 in response to microbial TLR ligands. After emerging from the germinal center, high-affinity IgA class-switched B cells enter the circulation and migrate to the lamina propria of the intestine, where they differentiate into long-lived IgA-secreting plasma cells.
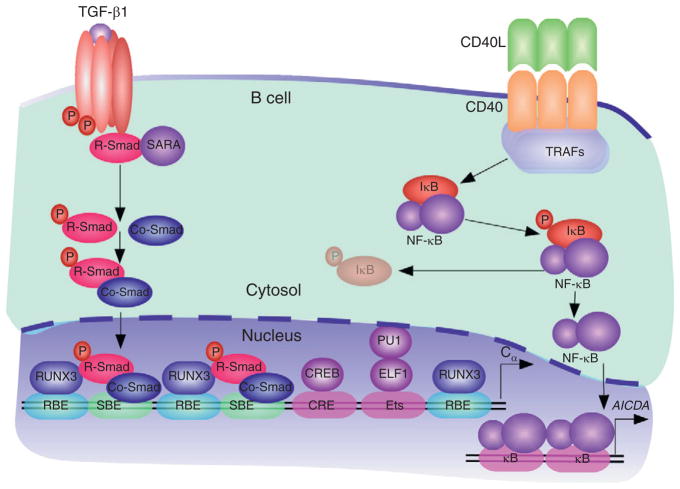
Signaling events in TD IgA CSR and production. TGF-β1 from T cells, DCs, FDCs, IECs, macrophages, and stromal cells forms a heteromeric TGF receptor (TGFR) complex on B cells that activates SMAD proteins. In the presence of TGF-β1, TGFβRII kinases phosphorylate TGFβRI, leading to the activation of TGFβRI kinases. These kinases induce the phosphorylation of receptor-regulated SMAD (R-SMAD) proteins, thereby releasing them from the plasma membrane-anchoring protein SARA (SMAD anchor for receptor activation). After forming homo-oligomeric complexes, as well as hetero-oligomeric complexes with a co-mediator SMAD (Co-SMAD) protein, R-SMAD proteins translocate to the nucleus, where they bind to SMAD-binding elements (SBEs) on target gene promoters, including constant heavy chain α (Cα) gene promoters. These SMAD complexes further associate with constitutive and TGFβR-induced cofactors, including RUNX3, which binds to RUNX-binding elements (RBEs), cyclic AMP response element-binding protein (CREB), which binds to a cyclic AMP response element (CRE), and Ets-like factor 1 (ELF1), which binds to an Ets-binding site together with PU.1. CD40L expressed on T cell surface elicits oligomerization of CD40 on B cells, recruitment of TRAFs to CD40, activation of the IKK complex, and phosphorylation and degradation of IκB. The resulting IκB-free NF-κB proteins translocate to the nucleus to induce transcription of the AICDA gene promoter and AID expression.
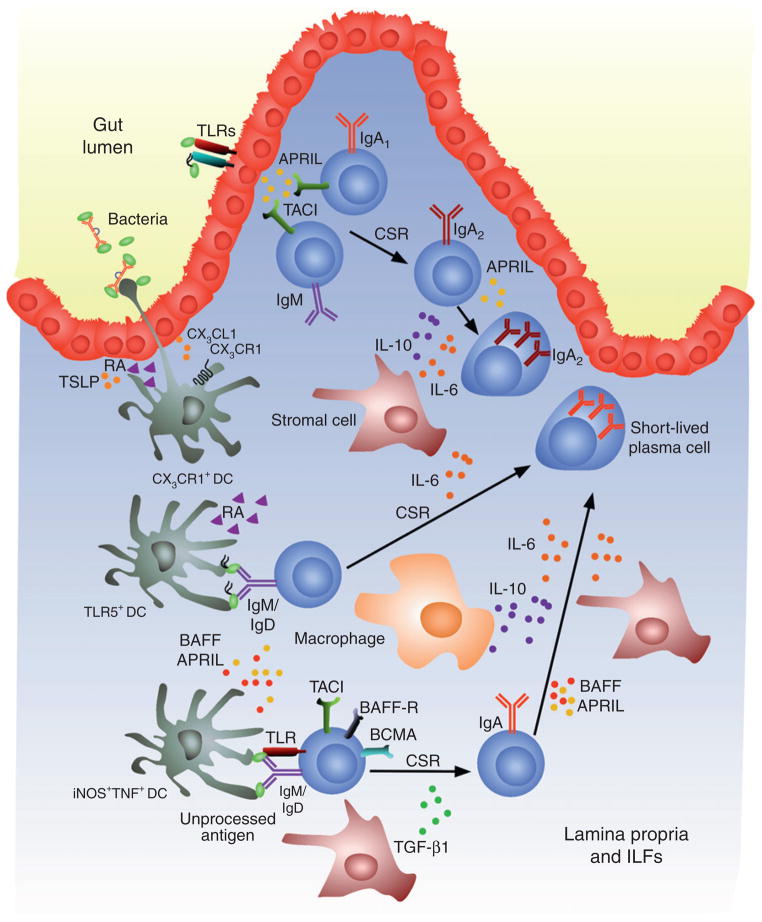
TI induction of IgA CSR and production in the intestinal lamina propria. IECs sense microorganism through TLRs and thereafter release APRIL, which triggers direct IgM-to-IgA1 CSR in lamina propria IgM+ B cells and sequential IgA1-to-IgA2 CSR in lamina propria IgA1+ B cells. TLR-activated IECs further amplify TI IgA CSR and production by stimulating DCs, including antigen-sampling CX3CR1+ DCs, iNOS+TNF+ DCs, and CD11chiCD11bhiTLR5+ DCs, through TSLP and RA. Together with stromal cells, macrophages, and lymphoid tissue-inducing cells (not shown), DCs would elicit IgA CSR and production in the nonorganized lymphoid tissue of the lamina propria or organized isolated lymphoid follicles (not shown) by releasing BAFF, APRIL, IL-6, IL-10, and TGF-β1, and by engaging BCR and TLRs through antigen. The resulting IgA class-switched B cells differentiate into short-lived IgA-secreting plasma cells.
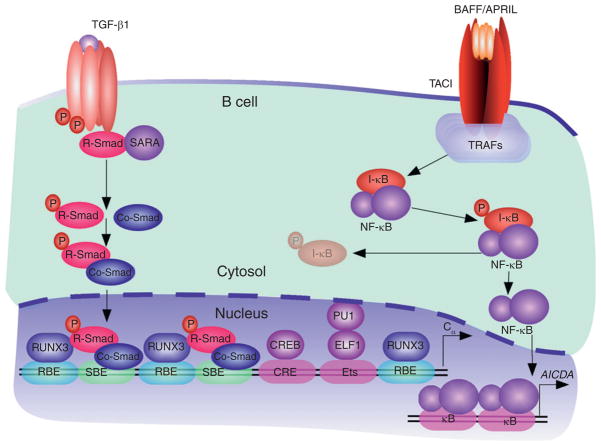
Signaling events in TI IgA CSR and production. TGF-β1 from T cells, DCs, FDCs, IECs, macrophages, and stromal cells forms a heteromeric TGF receptor (TGFR) complex on B cells that activates SMAD proteins. In the presence of TGF-β1, TGFβRII kinases phosphorylate TGFβRI, leading to the activation of TGFβRI kinases. These kinases induce the phosphorylation of receptor-regulated SMAD (R-SMAD) proteins, thereby releasing them from the plasma membrane-anchoring protein SARA (SMAD anchor for receptor activation). After forming homo-oligomeric complexes, as well as hetero-oligomeric complexes with a co-mediator SMAD (Co-SMAD) protein, R-SMAD proteins translocate to the nucleus, where they bind to SMAD-binding elements (SBEs) on target gene promoters, including Cα gene promoters. These SMAD complexes further associate with constitutive and TGFβR-induced cofactors, including RUNX3, which binds to RUNX-binding elements (RBEs), cyclic AMP response element-binding protein (CREB), which binds to a cyclic AMP response element (CRE), and Ets-like factor 1 (ELF1), which binds to an Ets-binding site together with PU.1. BAFF and APRIL from IECs, DCs, FDCs, macrophages, and stromal cells elicit oligomerization of TACI on B cells, recruitment of TRAFs to TACI, activation of the IKK complex, and phosphorylation and degradation of IκB. The resulting IκB-free NF-κB proteins translocate to the nucleus to induce transcription of the AICDA gene promoter and AID expression.
References
-
- Abreu MT. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat Rev Immunol. 2010;10:131–144. - PubMed
-
- Artis D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat Rev Immunol. 2008;8:411–420. - PubMed
-
- Avery DT, Deenick EK, Ma CS, Suryani S, Simpson N, Chew GY, Chan TD, Palendira U, Bustamante J, Boisson-Dupuis S, Choo S, Bleasel KE, et al. B cell-intrinsic signaling through IL-21 receptor and STAT3 is required for establishing long-lived antibody responses in humans. J Exp Med. 2010;207:155–171. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
