

Regulation of the histone H4 monomethylase PR-Set7 by CRL4(Cdt2)-mediated PCNA-dependent degradation during DNA damage
- PMID: 21035370
- PMCID: PMC2999913
- DOI: 10.1016/j.molcel.2010.10.011
Regulation of the histone H4 monomethylase PR-Set7 by CRL4(Cdt2)-mediated PCNA-dependent degradation during DNA damage
Abstract
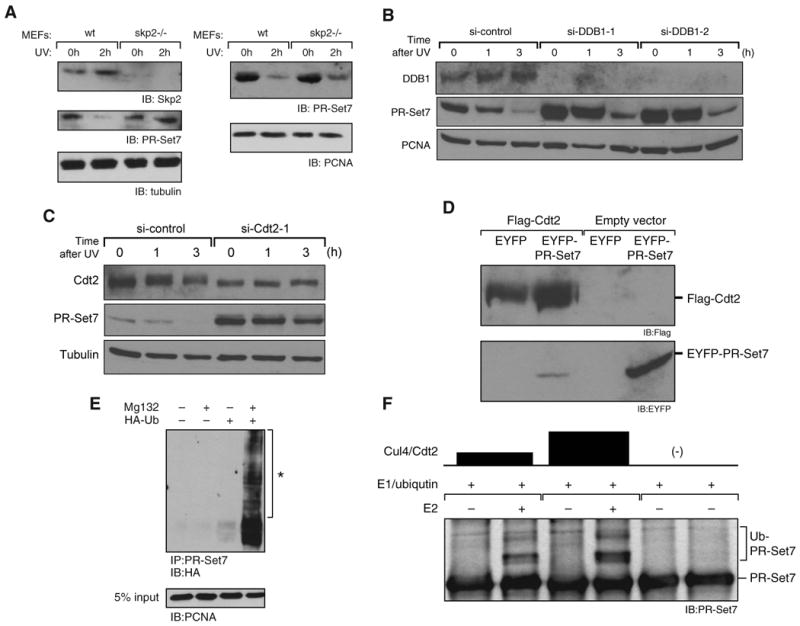
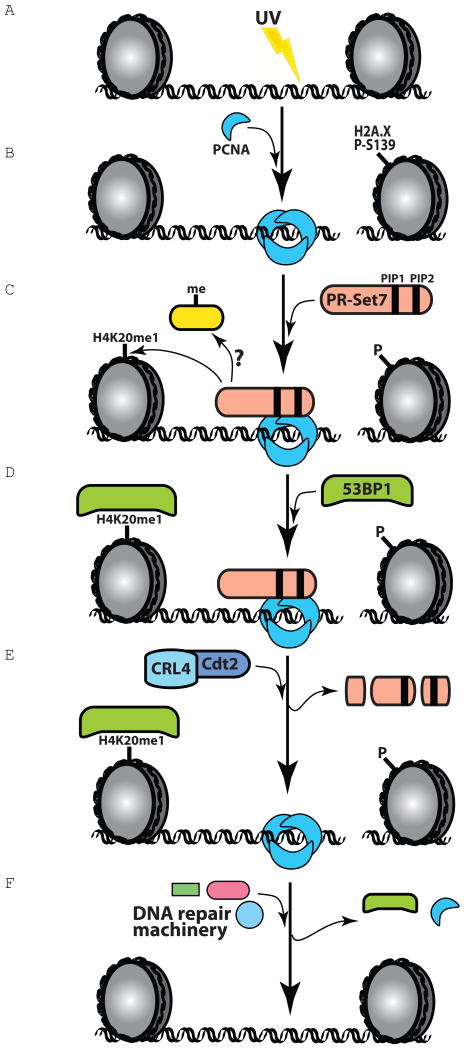
The histone methyltransferase PR-Set7/Set8 is the sole enzyme that catalyzes monomethylation of histone H4 at K20 (H4K20me1). Previous reports document disparate evidence regarding PR-Set7 expression during the cell cycle, the biological relevance of PR-Set7 interaction with PCNA, and its role in the cell. We find that PR-Set7 is indeed undetectable during S phase and instead is detected during late G2, mitosis, and early G1. PR-Set7 is transiently recruited to laser-induced DNA damage sites through its interaction with PCNA, after which 53BP1 is recruited dependent on PR-Set7 catalytic activity. During the DNA damage response, PR-Set7 interaction with PCNA through a specialized "PIP degron" domain targets it for PCNA-coupled CRL4(Cdt2)-dependent proteolysis. PR-Set7 mutant in its "PIP degron" is now detectable during S phase, during which the mutant protein accumulates. Outside the chromatin context, Skp2 promotes PR-Set7 degradation as well. These findings demonstrate a stringent spatiotemporal control of PR-Set7 that is essential for preserving the genomic integrity of mammalian cells.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
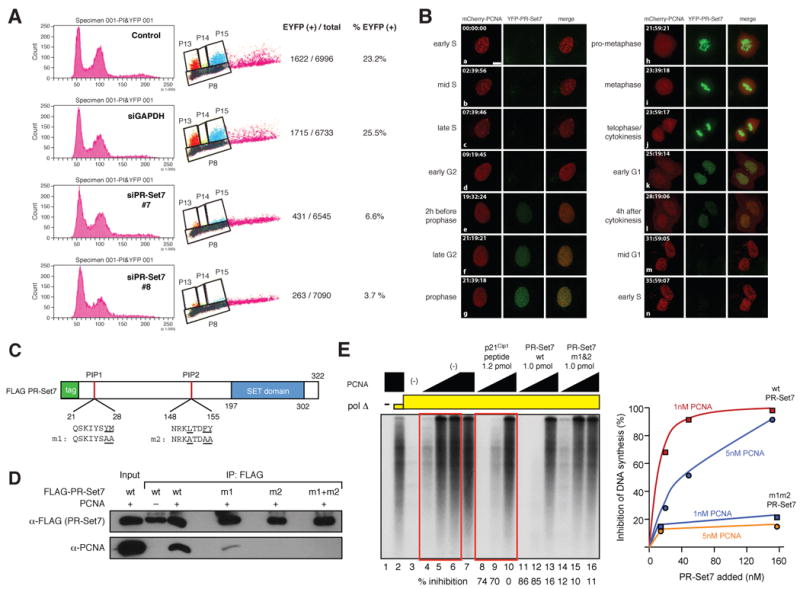
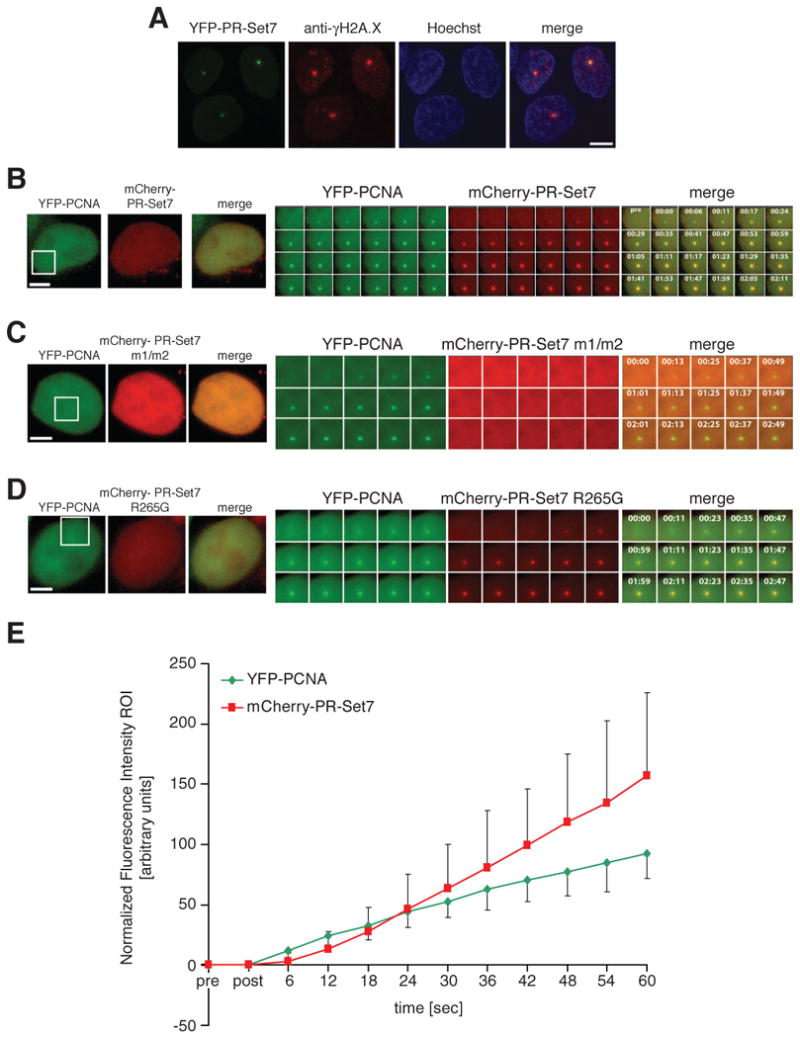
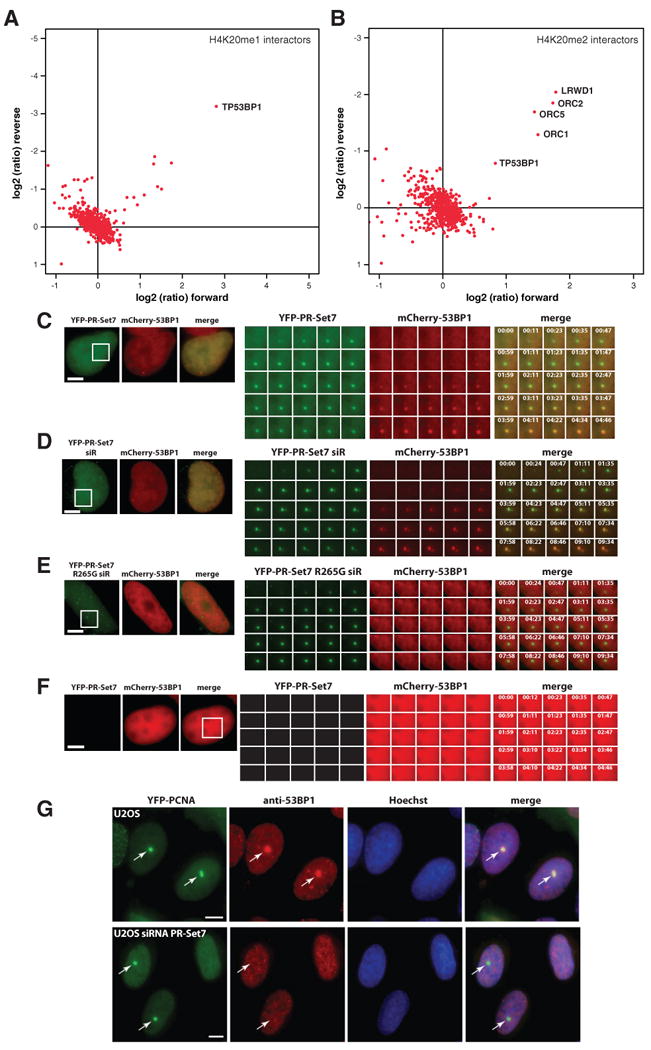
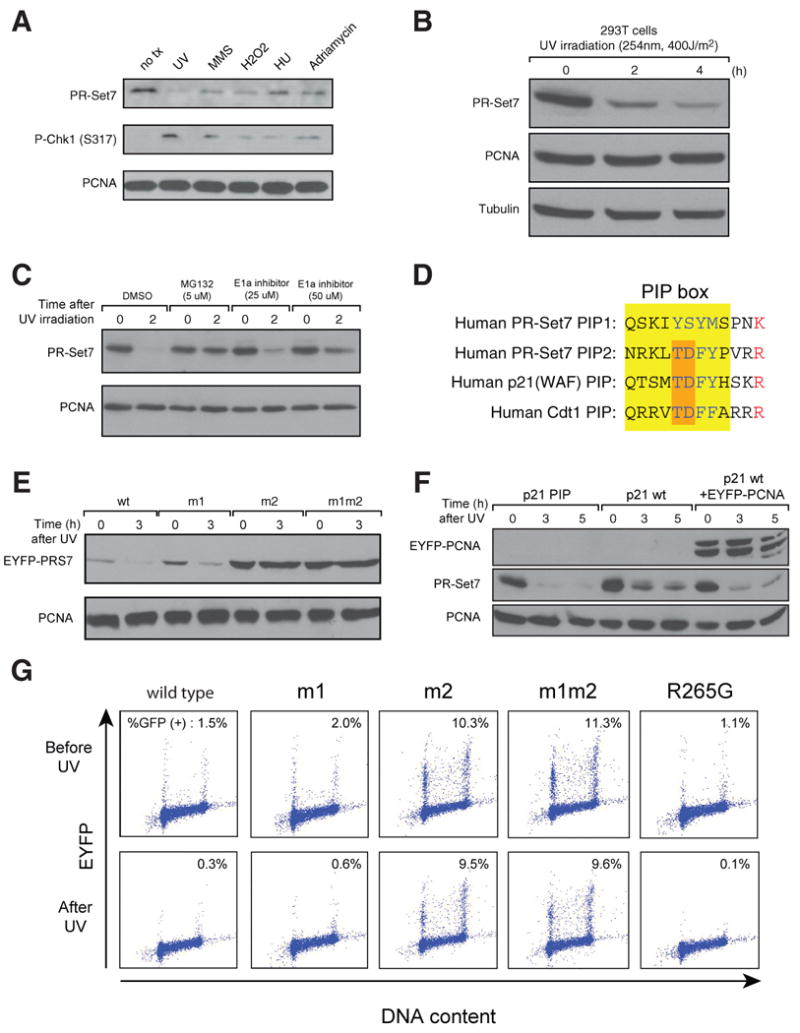
Comment in
-
SETting the clock for histone H4 monomethylation.Mol Cell. 2010 Nov 12;40(3):345-6. doi: 10.1016/j.molcel.2010.10.033. Mol Cell. 2010. PMID: 21070959 Free PMC article.
-
Cell cycle: Disposing of SETD8.Nat Rev Mol Cell Biol. 2010 Dec;11(12):819. doi: 10.1038/nrm3020. Nat Rev Mol Cell Biol. 2010. PMID: 21102605 No abstract available.
References
-
- Anastassiadis K, Kim J, Daigle N, Sprengel R, Scholer HR, Stewart AF. A predictable ligand regulated expression strategy for stably integrated transgenes in mammalian cells in culture. Gene. 2002;298:159–172. - PubMed
-
- Arias EE, Walter JC. PCNA functions as a molecular platform to trigger Cdt1 destruction and prevent re-replication. Nat Cell Biol. 2006;8:84–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
