

Crystal structure of the phage T4 recombinase UvsX and its functional interaction with the T4 SF2 helicase UvsW
- PMID: 21035462
- PMCID: PMC3006652
- DOI: 10.1016/j.jmb.2010.10.004
Crystal structure of the phage T4 recombinase UvsX and its functional interaction with the T4 SF2 helicase UvsW
Abstract
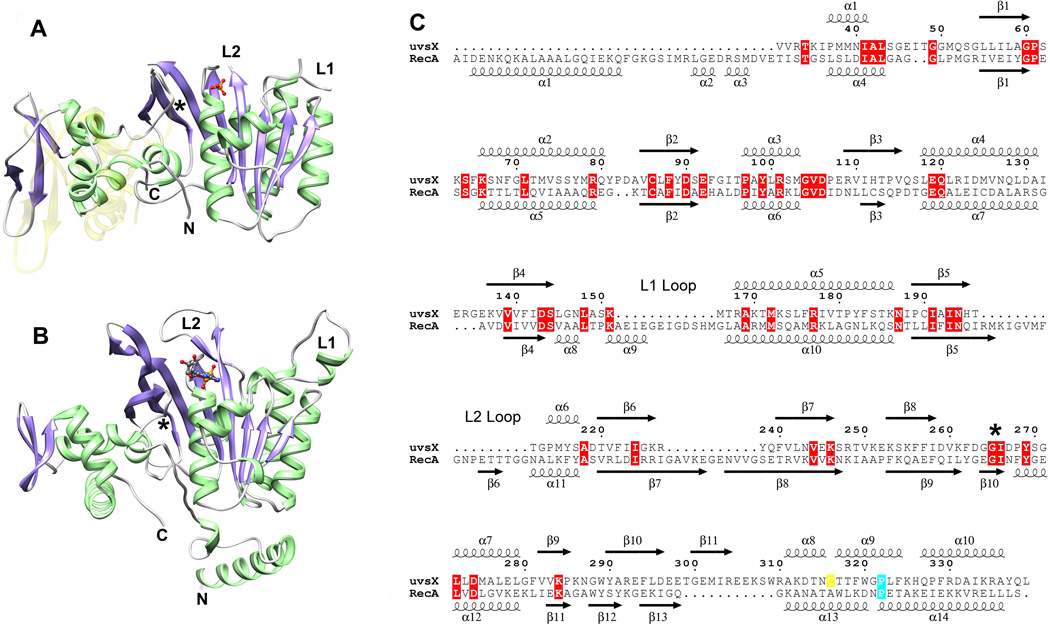
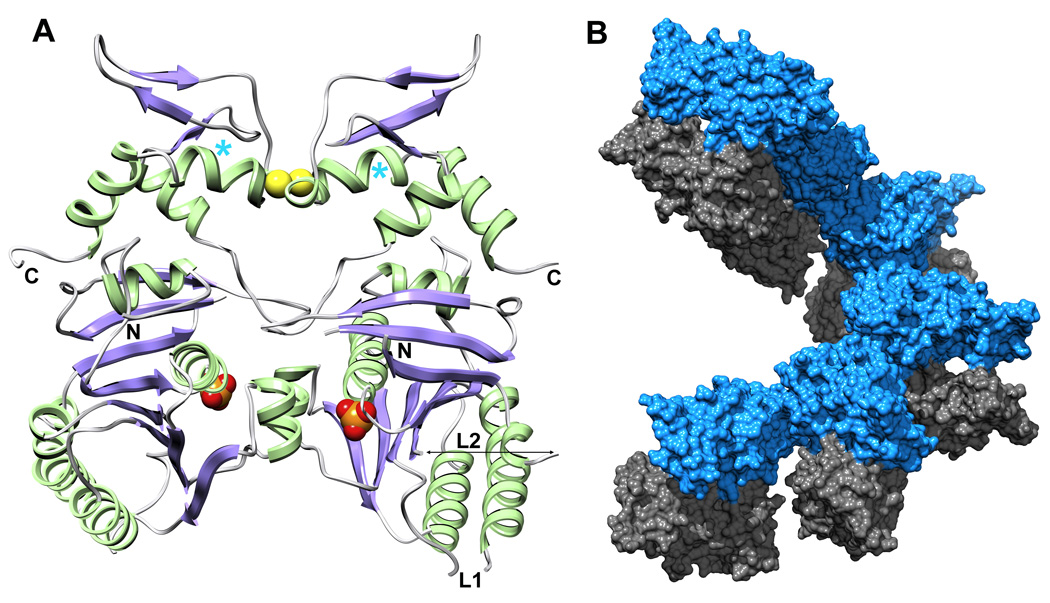
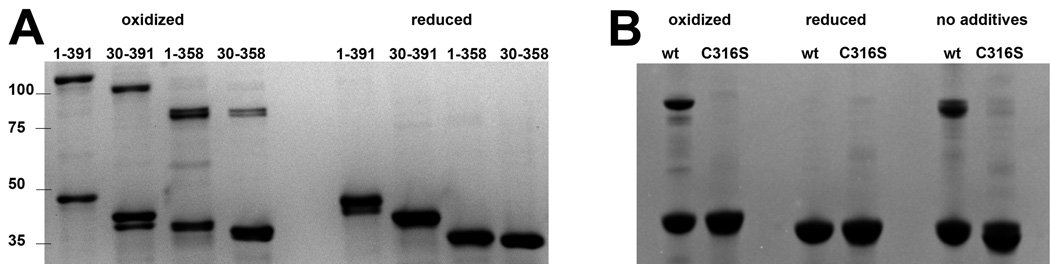
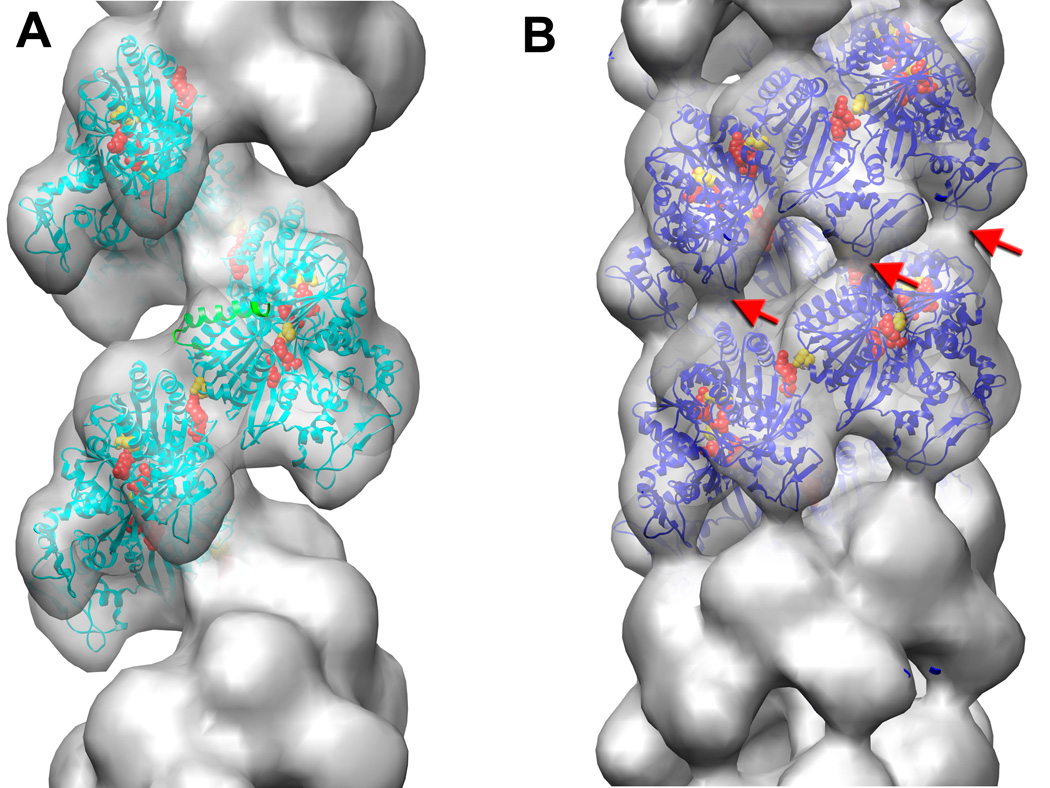
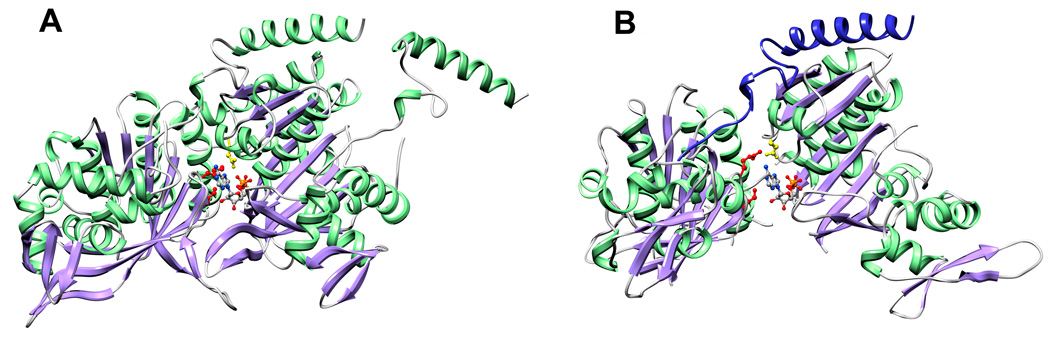
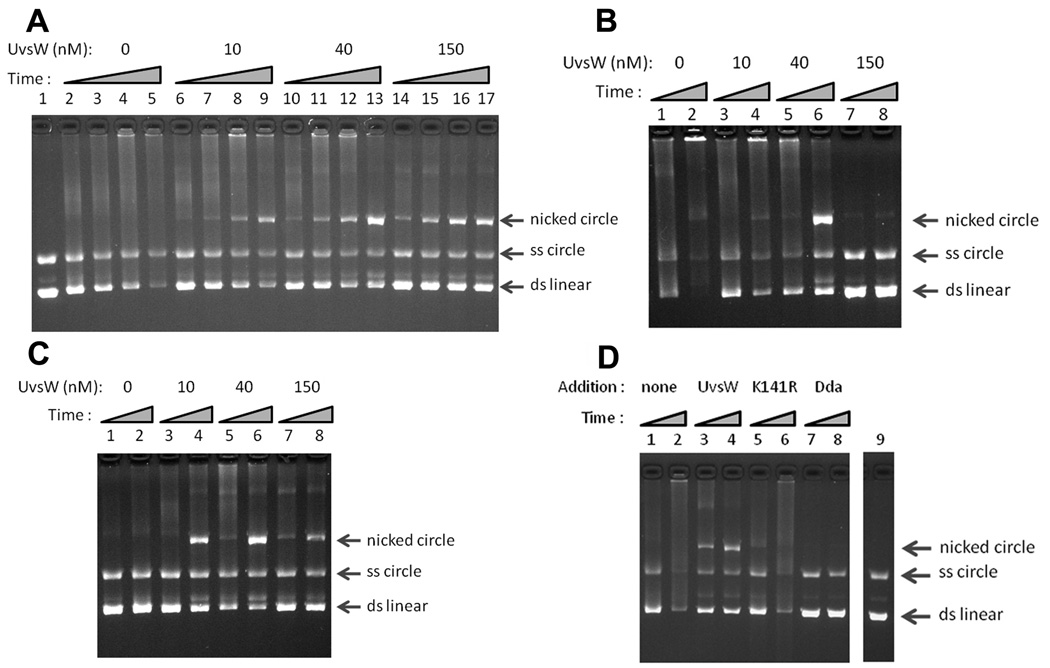
Bacteriophage T4 provides an important model system for studying the mechanism of homologous recombination. We have determined the crystal structure of the T4 UvsX recombinase, and the overall architecture and fold closely resemble those of RecA, including a highly conserved ATP binding site. Based on this new structure, we reanalyzed electron microscopy reconstructions of UvsX-DNA filaments and docked the UvsX crystal structure into two different filament forms: a compressed filament generated in the presence of ADP and an elongated filament generated in the presence of ATP and aluminum fluoride. In these reconstructions, the ATP binding site sits at the protomer interface, as in the RecA filament crystal structure. However, the environment of the ATP binding site is altered in the two filament reconstructions, suggesting that nucleotide cannot be as easily accommodated at the protomer interface of the compressed filament. Finally, we show that the phage helicase UvsW completes the UvsX-promoted strand-exchange reaction, allowing the generation of a simple nicked circular product rather than complex networks of partially exchanged substrates.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Comparison of bacteriophage T4 UvsX and human Rad51 filaments suggests that RecA-like polymers may have evolved independently.J Mol Biol. 2001 Oct 5;312(5):999-1009. doi: 10.1006/jmbi.2001.5025. J Mol Biol. 2001. PMID: 11580245
-
The crystal structure of the UvsW helicase from bacteriophage T4.Structure. 2004 Apr;12(4):583-92. doi: 10.1016/j.str.2004.02.016. Structure. 2004. PMID: 15062081
-
Role of allosteric switch residue histidine 195 in maintaining active-site asymmetry in presynaptic filaments of bacteriophage T4 UvsX recombinase.J Mol Biol. 2009 Jan 16;385(2):393-404. doi: 10.1016/j.jmb.2008.11.003. Epub 2008 Nov 12. J Mol Biol. 2009. PMID: 19027026 Free PMC article.
-
Mediator proteins orchestrate enzyme-ssDNA assembly during T4 recombination-dependent DNA replication and repair.Proc Natl Acad Sci U S A. 2001 Jul 17;98(15):8298-305. doi: 10.1073/pnas.131007498. Proc Natl Acad Sci U S A. 2001. PMID: 11459967 Free PMC article. Review.
-
Assembly and dynamics of the bacteriophage T4 homologous recombination machinery.Virol J. 2010 Dec 3;7:357. doi: 10.1186/1743-422X-7-357. Virol J. 2010. PMID: 21129202 Free PMC article. Review.
Cited by
-
DNA-pairing and annealing processes in homologous recombination and homology-directed repair.Cold Spring Harb Perspect Biol. 2015 Feb 2;7(2):a016444. doi: 10.1101/cshperspect.a016444. Cold Spring Harb Perspect Biol. 2015. PMID: 25646379 Free PMC article. Review.
-
Coordination and processing of DNA ends during double-strand break repair: the role of the bacteriophage T4 Mre11/Rad50 (MR) complex.Genetics. 2013 Nov;195(3):739-55. doi: 10.1534/genetics.113.154872. Epub 2013 Aug 26. Genetics. 2013. PMID: 23979587 Free PMC article.
-
Coordinated Binding of Single-Stranded and Double-Stranded DNA by UvsX Recombinase.PLoS One. 2013 Jun 18;8(6):e66654. doi: 10.1371/journal.pone.0066654. Print 2013. PLoS One. 2013. PMID: 23824136 Free PMC article.
-
Computational Insights into the Dynamic Structural Features and Binding Characteristics of Recombinase UvsX Compared with RecA.Molecules. 2023 Apr 11;28(8):3363. doi: 10.3390/molecules28083363. Molecules. 2023. PMID: 37110596 Free PMC article.
-
Nucleoside Triphosphate Phosphohydrolase I (NPH I) Functions as a 5' to 3' Translocase in Transcription Termination of Vaccinia Early Genes.J Biol Chem. 2016 Jul 8;291(28):14826-38. doi: 10.1074/jbc.M116.730135. Epub 2016 May 6. J Biol Chem. 2016. PMID: 27189950 Free PMC article.
References
-
- Bianco PR, Tracy RB, Kowalczykowski SC. DNA strand exchange proteins: a biochemical and physical comparison. Front Biosci. 1998;3:D570–D603. - PubMed
-
- West SC. Molecular views of recombination proteins and their control. Nat Rev Mol Cell Biol. 2003;4:435–445. - PubMed
-
- Cox MM, Goodman MF, Kreuzer KN, Sherratt DJ, Sandler SJ, Marians KJ. The importance of repairing stalled replication forks. Nature. 2000;404:37–41. - PubMed
-
- San Filippo J, Sung P, Klein H. Mechanism of eukaryotic homologous recombination. Annu Rev Biochem. 2008;77:229–257. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
