

Complementary synaptic distribution of enzymes responsible for synthesis and inactivation of the endocannabinoid 2-arachidonoylglycerol in the human hippocampus
- PMID: 21035522
- PMCID: PMC3678284
- DOI: 10.1016/j.neuroscience.2010.10.062
Complementary synaptic distribution of enzymes responsible for synthesis and inactivation of the endocannabinoid 2-arachidonoylglycerol in the human hippocampus
Erratum in
- Neuroscience. 2011 Jul 14;186:220
Abstract
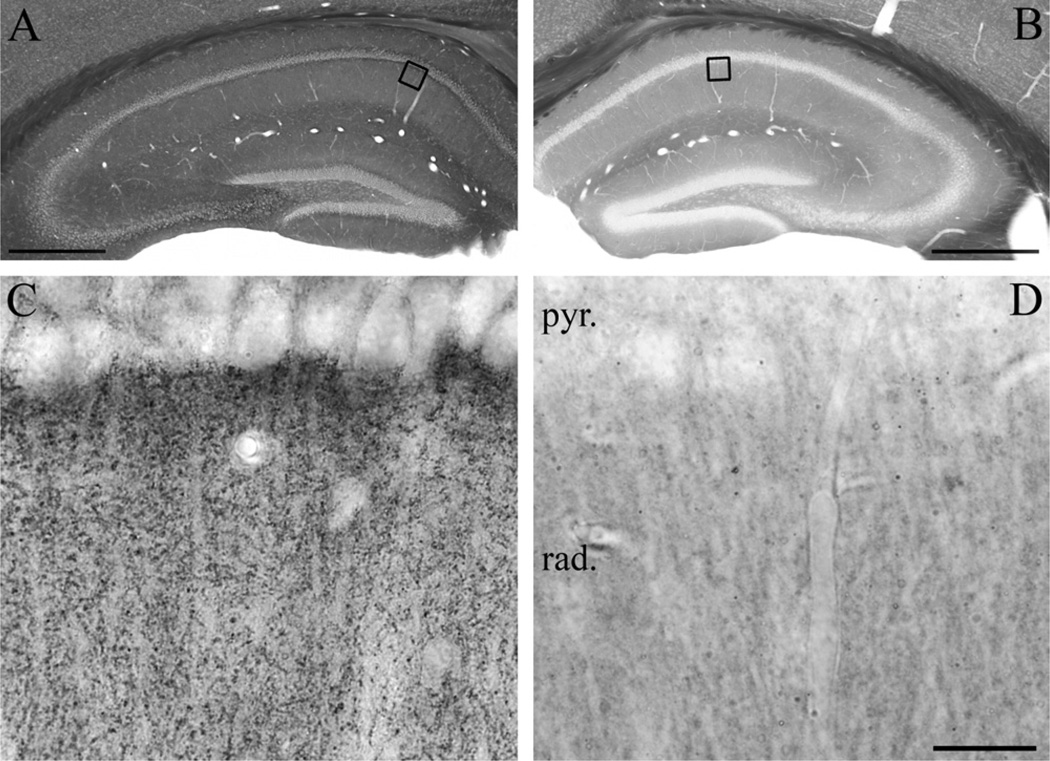
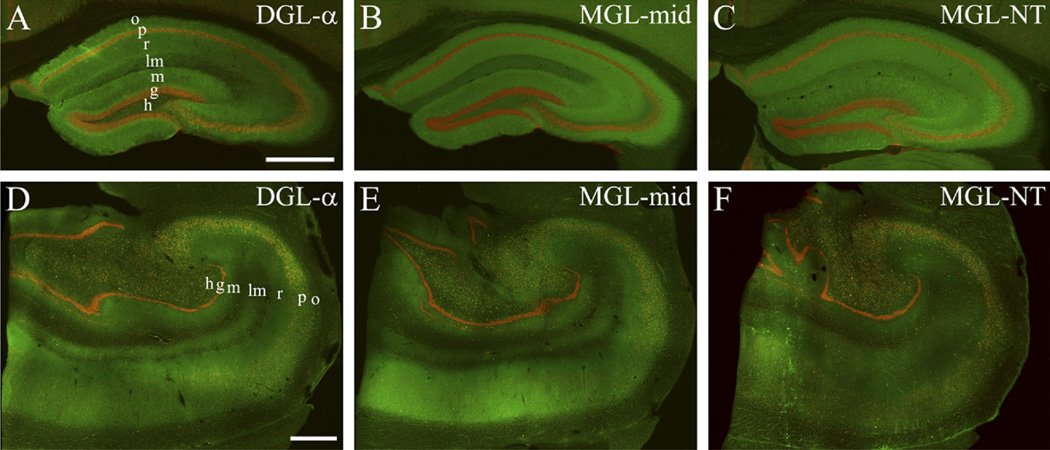
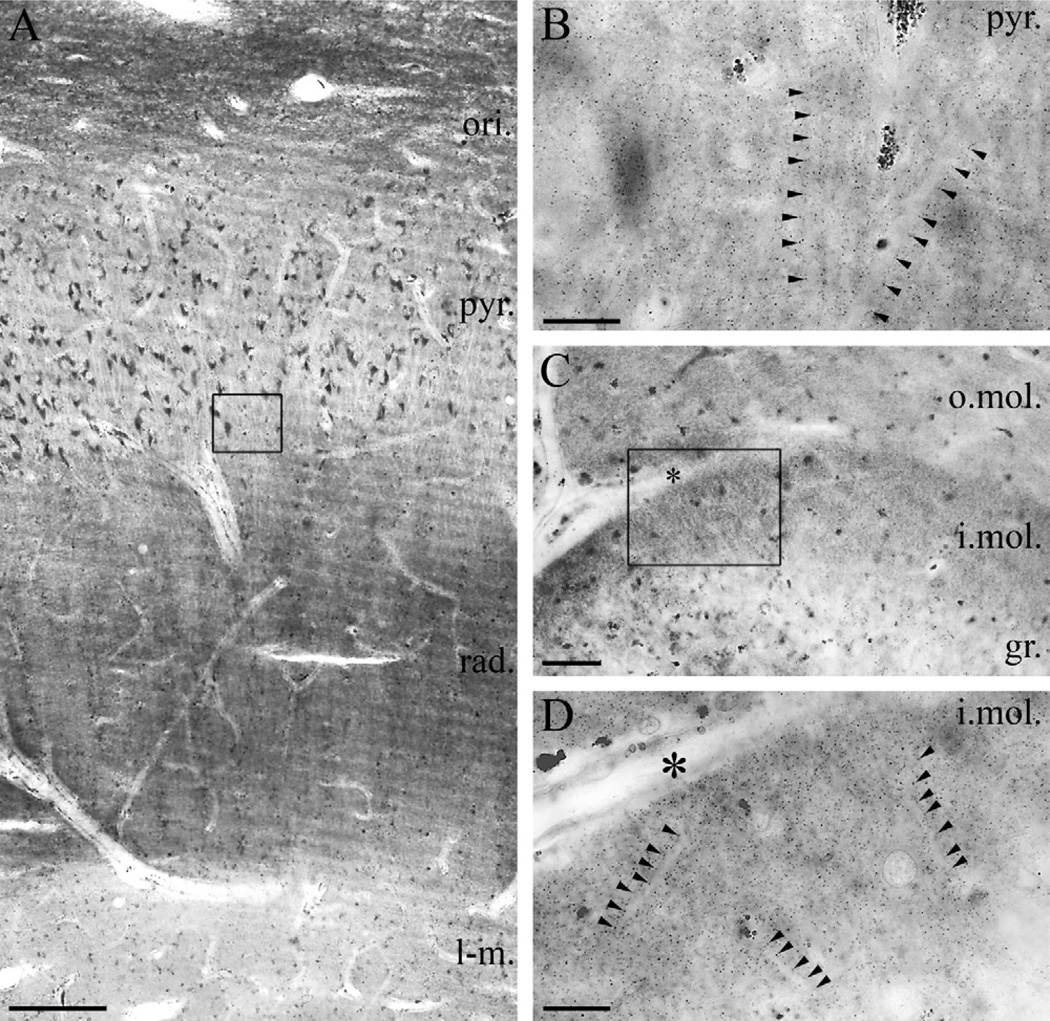
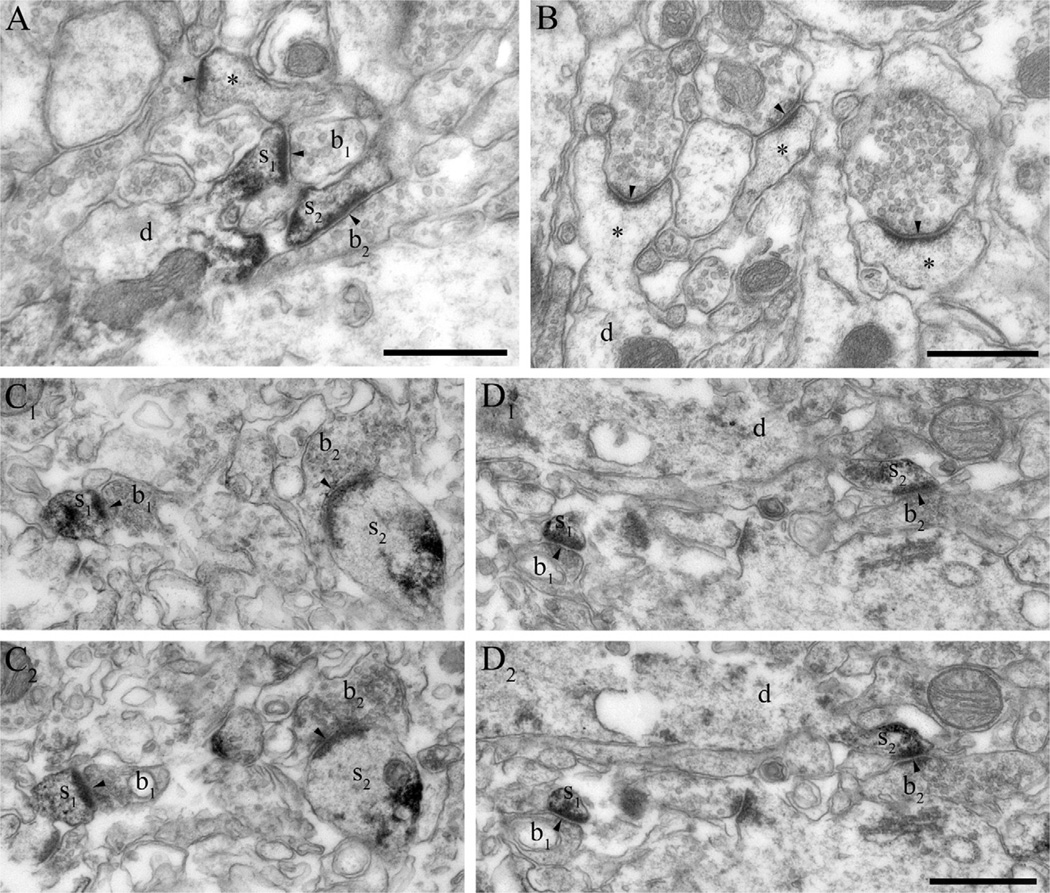
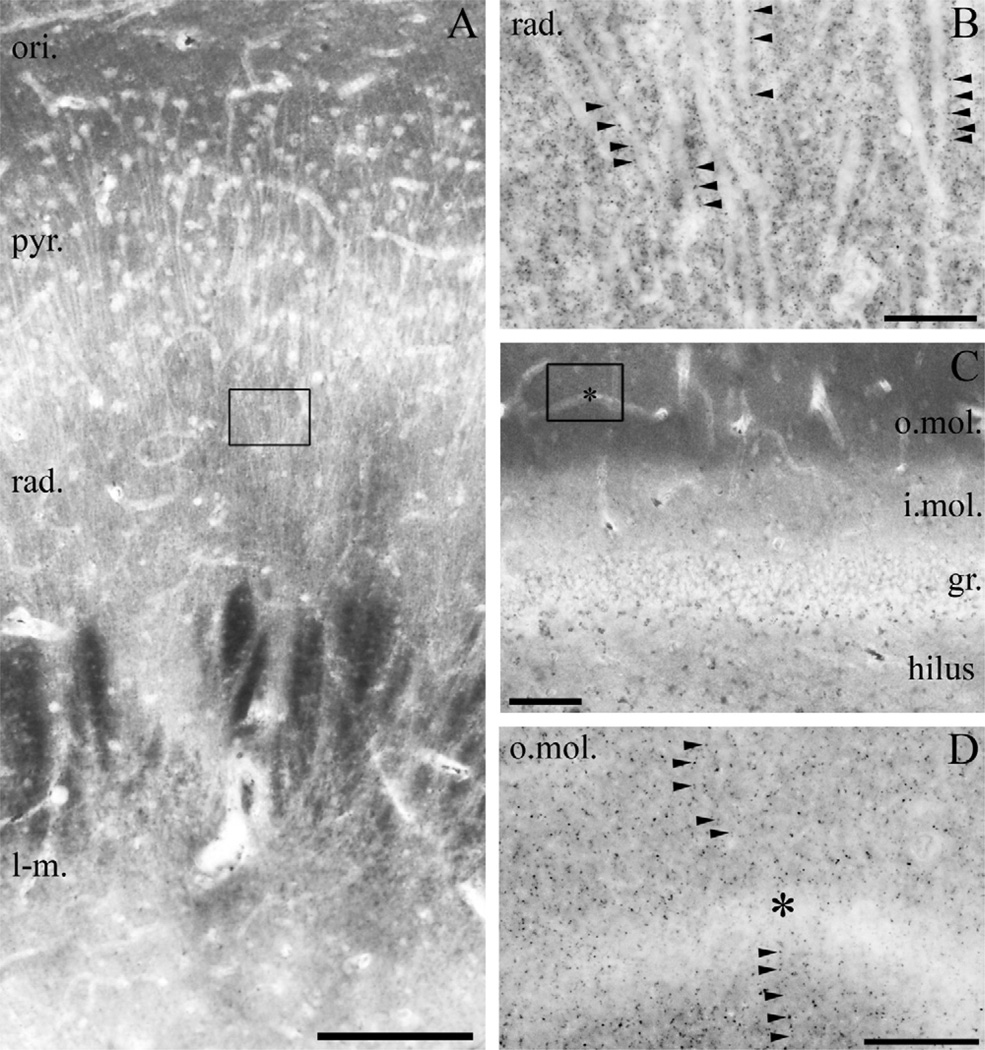
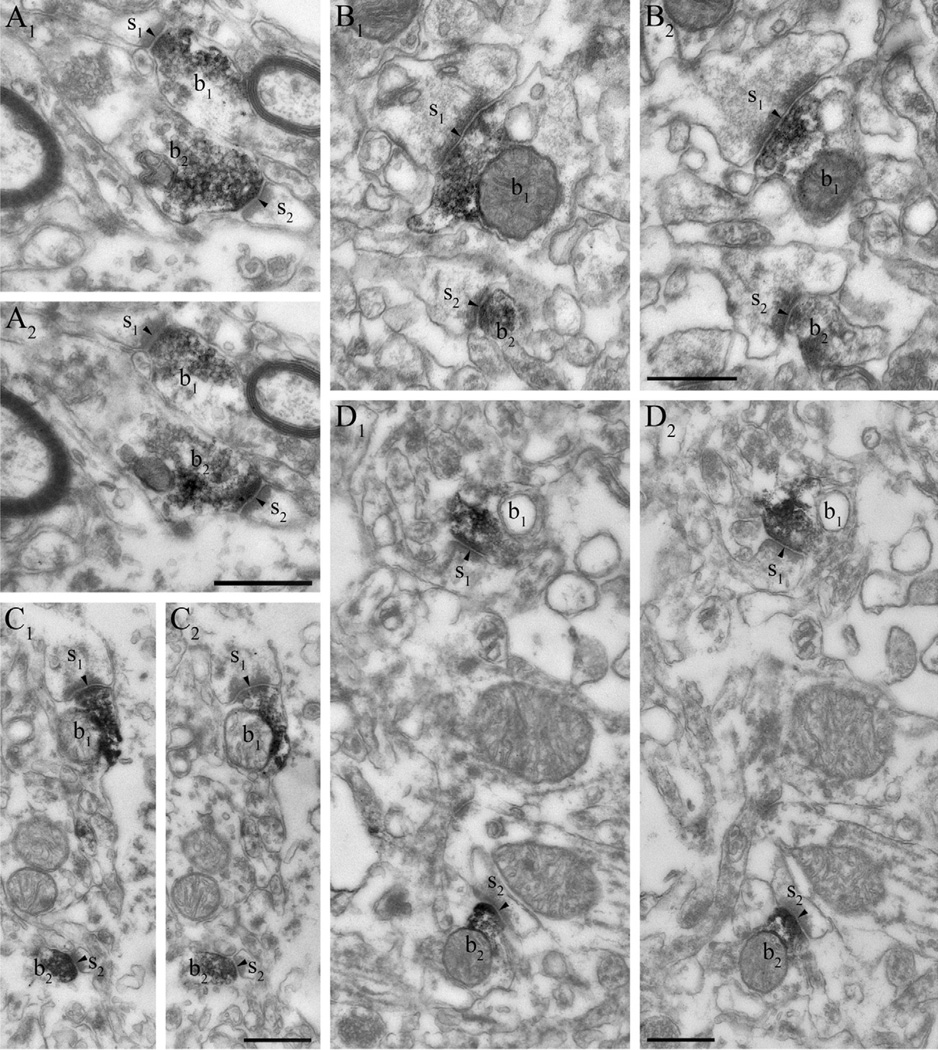
Clinical and experimental evidence demonstrates that endocannabinoids play either beneficial or adverse roles in many neurological and psychiatric disorders. Their medical significance may be best explained by the emerging concept that endocannabinoids are essential modulators of synaptic transmission throughout the central nervous system. However, the precise molecular architecture of the endocannabinoid signaling machinery in the human brain remains elusive. To address this issue, we investigated the synaptic distribution of metabolic enzymes for the most abundant endocannabinoid molecule, 2-arachidonoylglycerol (2-AG), in the postmortem human hippocampus. Immunostaining for diacylglycerol lipase-α (DGL-α), the main synthesizing enzyme of 2-AG, resulted in a laminar pattern corresponding to the termination zones of glutamatergic pathways. The highest density of DGL-α-immunostaining was observed in strata radiatum and oriens of the cornu ammonis and in the inner third of stratum moleculare of the dentate gyrus. At higher magnification, DGL-α-immunopositive puncta were distributed throughout the neuropil outlining the immunonegative main dendrites of pyramidal and granule cells. Electron microscopic analysis revealed that this pattern was due to the accumulation of DGL-α in dendritic spine heads. Similar DGL-α-immunostaining pattern was also found in hippocampi of wild-type, but not of DGL-α knockout mice. Using two independent antibodies developed against monoacylglycerol lipase (MGL), the predominant enzyme inactivating 2-AG, immunostaining also revealed a laminar and punctate staining pattern. However, as observed previously in rodent hippocampus, MGL was enriched in axon terminals instead of postsynaptic structures at the ultrastructural level. Taken together, these findings demonstrate the post- and presynaptic segregation of primary enzymes responsible for synthesis and elimination of 2-AG, respectively, in the human hippocampus. Thus, molecular architecture of the endocannabinoid signaling machinery supports retrograde regulation of synaptic activity, and its similar blueprint in rodents and humans further indicates that 2-AG's physiological role as a negative feed-back signal is an evolutionarily conserved feature of excitatory synapses.
Copyright © 2011 IBRO. All rights reserved.
Figures
References
-
- Ahn K, Johnson DS, Mileni M, Beidler D, Long JZ, McKinney MK, Weerapana E, Sadagopan N, Liimatta M, Smith SE, Lazerwith S, Stiff C, Kamtekar S, Bhattacharya K, Zhang Y, Swaney S, Van Becelaere K, Stevens RC, Cravatt BF. Discovery and characterization of a highly selective FAAH inhibitor that reduces inflammatory pain. Chem Biol. 2009;16:411–420. - PMC - PubMed
-
- Bisogno T, Burston JJ, Rai R, Allara M, Saha B, Mahadevan A, Razdan RK, Wiley JL, Di Marzo V. Synthesis and pharmacological activity of a potent inhibitor of the biosynthesis of the endocannabinoid 2-arachidonoylglycerol. Chem Med Chem. 2009;4:946–950. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MH54671/MH/NIMH NIH HHS/United States
- DA011322/DA/NIDA NIH HHS/United States
- R01 MH054671/MH/NIMH NIH HHS/United States
- R01 NS030549/NS/NINDS NIH HHS/United States
- DA021696/DA/NIDA NIH HHS/United States
- R37 NS030549/NS/NINDS NIH HHS/United States
- R01 DA011322/DA/NIDA NIH HHS/United States
- NS030549/NS/NINDS NIH HHS/United States
- 243153/ERC_/European Research Council/International
- P01 DA009158/DA/NIDA NIH HHS/United States
- K05 DA021696/DA/NIDA NIH HHS/United States
- DA09158/DA/NIDA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
