

Bryostatin-1, a naturally occurring antineoplastic agent, acts as a Toll-like receptor 4 (TLR-4) ligand and induces unique cytokines and chemokines in dendritic cells
- PMID: 21036898
- PMCID: PMC3012980
- DOI: 10.1074/jbc.M110.135921
Bryostatin-1, a naturally occurring antineoplastic agent, acts as a Toll-like receptor 4 (TLR-4) ligand and induces unique cytokines and chemokines in dendritic cells
Abstract
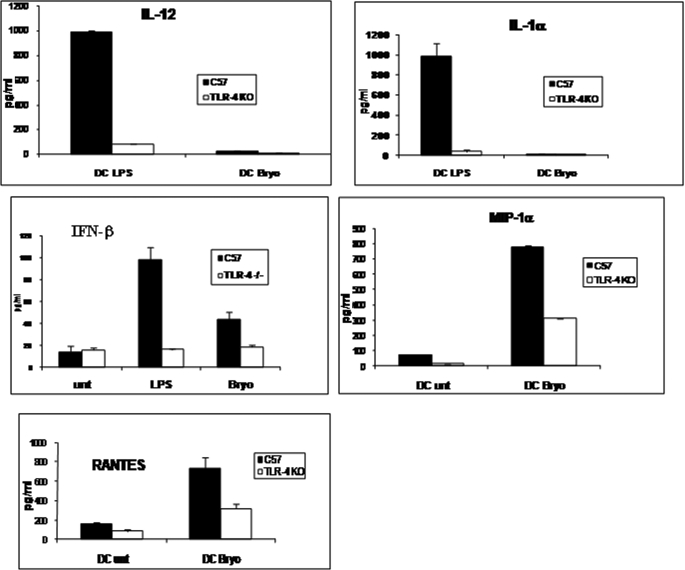
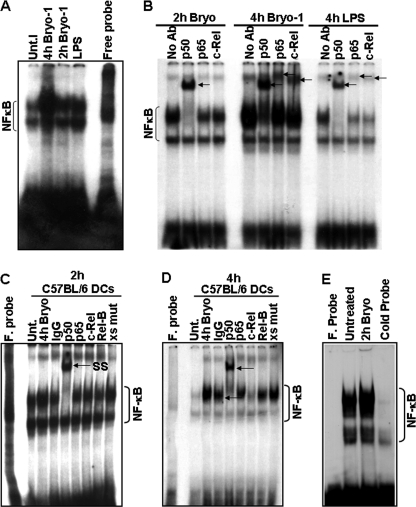
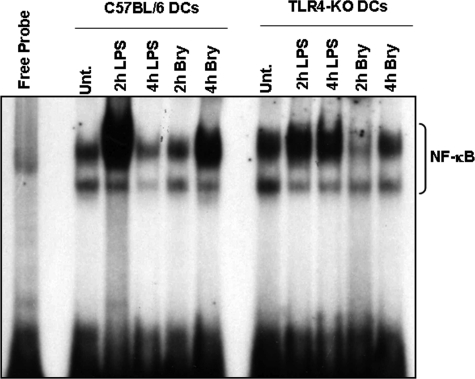
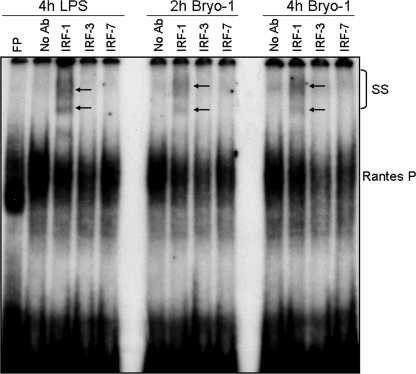
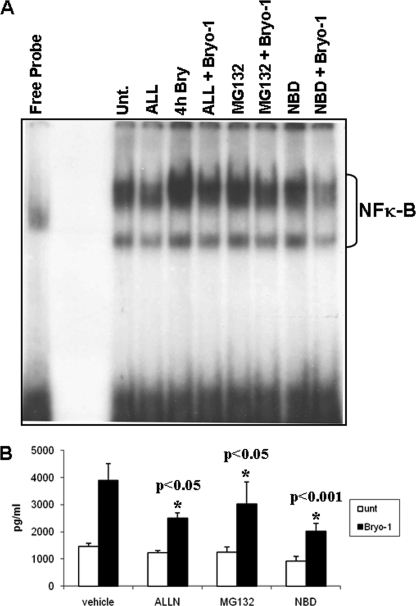
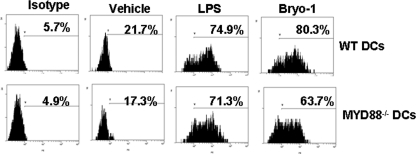
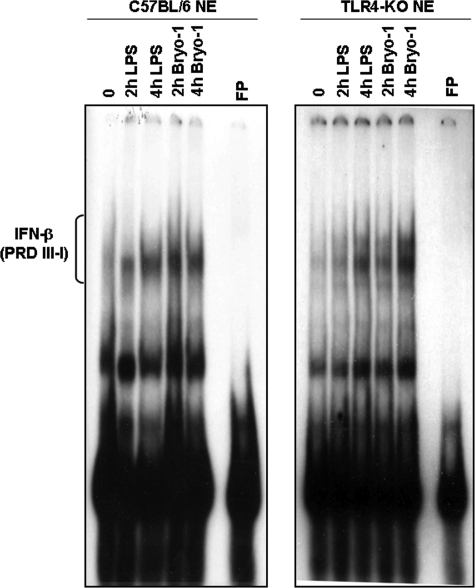
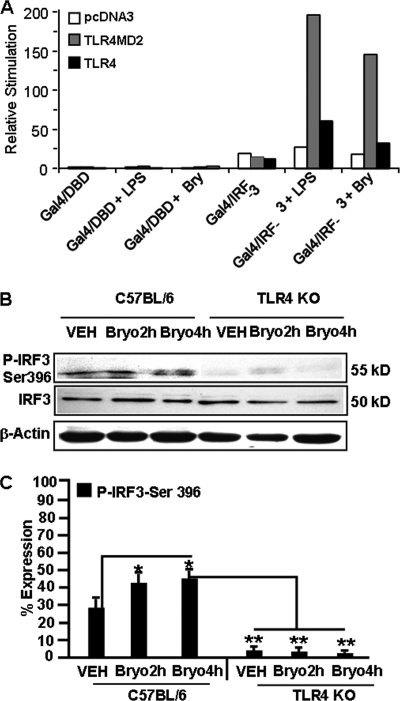
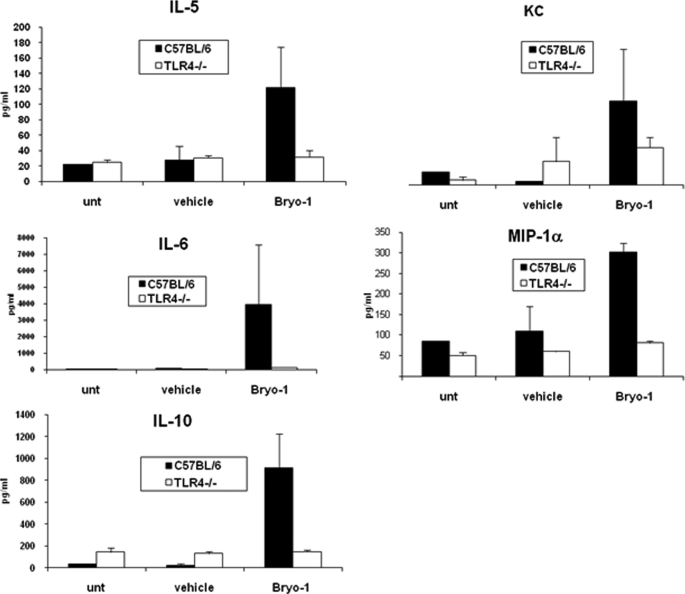
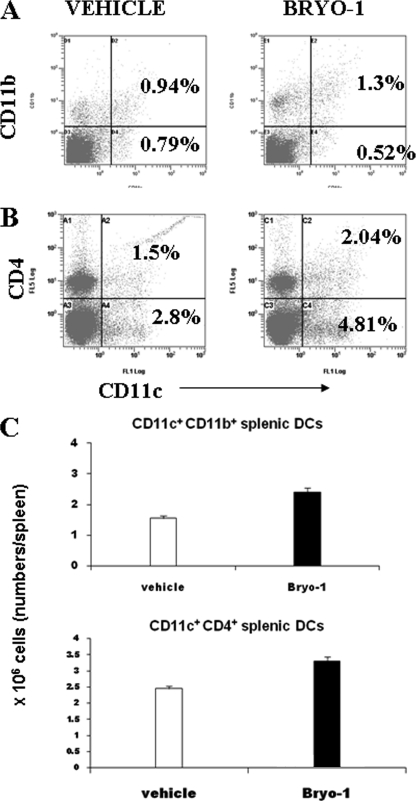
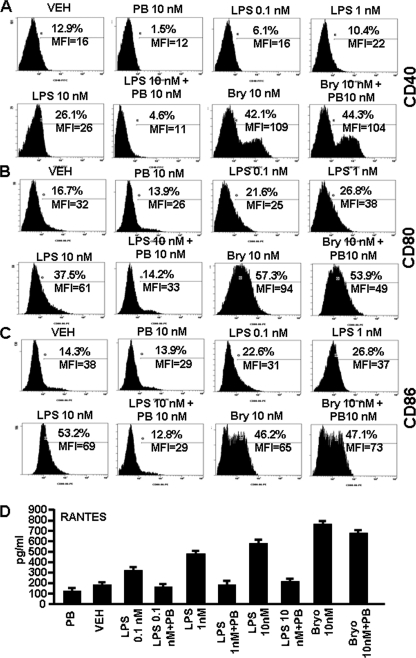
Bryostatin-1 (Bryo-1), a natural macrocyclic lactone, is clinically used as an anti-cancer agent. In this study, we demonstrate for the first time that Bryo-1 acts as a Toll-like receptor 4 (TLR4) ligand. Interestingly, activation of bone marrow-derived dendritic cells (in vitro with Bryo-1) led to a TLR4-dependent biphasic activation of nuclear factor-κB (NF-κB) and the unique induction of cytokines (IL-5, IL-6, and IL-10) and chemokines, including RANTES (regulated on activation normal T cell expressed and secreted) and macrophage inflammatory protein 1α (MIP1-α). In addition, EMSA demonstrated that Bryo-1-mediated induction of RANTES was regulated by NF-κB and the interferon regulatory factors (IRF)-1, IRF-3, and IRF-7 to the RANTES independently of myeloid differentiation primary response gene-88 (MyD88). Bryo-1 was able to induce the transcriptional activation of IRF-3 through the TLR4/MD2-dependent pathway. In vivo administration of Bryo-1 triggered a TLR-4-dependent T helper cell 2 (Th2) cytokine response and expanded a subset of myeloid dendritic cells that expressed a CD11c(high)CD8α(-) CD11b(+)CD4(+) phenotype. This study demonstrates that Bryo-1 can act as a TLR4 ligand and activate innate immunity. Moreover, the ability of Bryo-1 to trigger RANTES and MIP1-α suggests that Bryo-1 could potentially be used to prevent HIV-1 infection. Finally, induction of a Th2 response by Bryo-1 may help treat inflammatory diseases mediated by Th1 cells. Together, our studies have a major impact on the clinical use of Bryo-1 as an anti-cancer and immunopotentiating agent.
Figures
References
-
- Montoya C. J., Jie H. B., Al-Harthi L., Mulder C., Patiño P. J., Rugeles M. T., Krieg A. M., Landay A. L., Wilson S. B. (2006) J. Immunol. 177, 1028–1039 - PubMed
-
- Banchereau J., Briere F., Caux C., Davoust J., Lebecque S., Liu Y. J., Pulendran B., Palucka K. (2000) Annu. Rev. Immunol. 18, 767–811 - PubMed
-
- Zenke M., Hieronymus T. (2006) Trends Immunol. 27, 140–145 - PubMed
-
- Medzhitov R., Janeway C., Jr. (2000) Trends Microbiol. 8, 452–456 - PubMed
-
- Kaisho T., Akira S. (2001) Trends Immunol. 22, 78–83 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
