

Dbf4 regulates the Cdc5 Polo-like kinase through a distinct non-canonical binding interaction
- PMID: 21036905
- PMCID: PMC3009850
- DOI: 10.1074/jbc.M110.155242
Dbf4 regulates the Cdc5 Polo-like kinase through a distinct non-canonical binding interaction
Abstract
Cdc7-Dbf4 is a conserved, two-subunit kinase required for initiating eukaryotic DNA replication. Recent studies have shown that Cdc7-Dbf4 also regulates the mitotic exit network (MEN) and monopolar homolog orientation in meiosis I (Matos, J., Lipp, J. J., Bogdanova, A., Guillot, S., Okaz, E., Junqueira, M., Shevchenko, A., and Zachariae, W. (2008) Cell 135, 662-678 and Miller, C. T., Gabrielse, C., Chen, Y. C., and Weinreich, M. (2009) PLoS Genet. 5, e1000498). Both activities likely involve a Cdc7-Dbf4 interaction with Cdc5, the single Polo-like kinase in budding yeast. We previously showed that Dbf4 binds the Cdc5 polo-box domain (PBD) via an ∼40-residue N-terminal sequence, which lacks a PBD consensus binding site (S(pS/pT)(P/X)), and that Dbf4 inhibits Cdc5 function during mitosis. Here we identify a non-consensus PBD binding site within Dbf4 and demonstrate that the PBD-Dbf4 interaction occurs via a distinct PBD surface from that used to bind phosphoproteins. Genetic and biochemical analysis of multiple dbf4 mutants indicate that Dbf4 inhibits Cdc5 function through direct binding. Surprisingly, mutation of invariant Cdc5 residues required for binding phosphorylated substrates has little effect on yeast viability or growth rate. Instead, cdc5 mutants defective for binding phosphoproteins exhibit enhanced resistance to microtubule disruption and an increased rate of spindle elongation. This study, therefore, details the molecular nature of a new type of PBD binding and reveals that Cdc5 targeting to phosphorylated substrates likely regulates spindle dynamics.
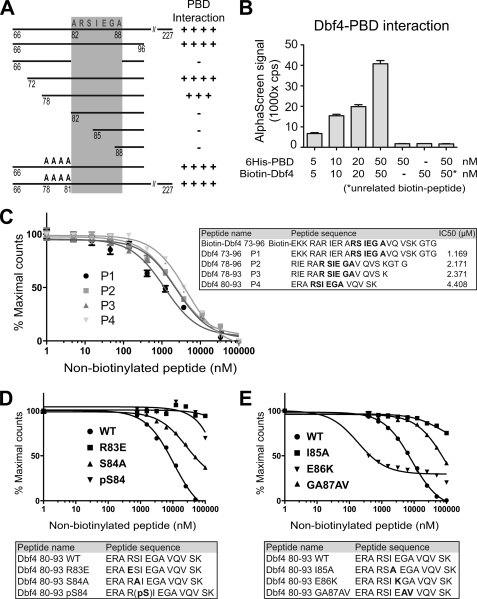
Figures





Similar articles
-
Dbf4: the whole is greater than the sum of its parts.Cell Cycle. 2013 Apr 15;12(8):1180-8. doi: 10.4161/cc.24416. Epub 2013 Apr 2. Cell Cycle. 2013. PMID: 23549174 Free PMC article. Review.
-
Distinct surfaces on Cdc5/PLK Polo-box domain orchestrate combinatorial substrate recognition during cell division.Sci Rep. 2020 Feb 25;10(1):3379. doi: 10.1038/s41598-020-60344-4. Sci Rep. 2020. PMID: 32099015 Free PMC article.
-
Cdc7p-Dbf4p regulates mitotic exit by inhibiting Polo kinase.PLoS Genet. 2009 May;5(5):e1000498. doi: 10.1371/journal.pgen.1000498. Epub 2009 May 29. PLoS Genet. 2009. PMID: 19478884 Free PMC article.
-
Independent modulation of the kinase and polo-box activities of Cdc5 protein unravels unique roles in the maintenance of genome stability.Proc Natl Acad Sci U S A. 2011 Oct 25;108(43):E914-23. doi: 10.1073/pnas.1106448108. Epub 2011 Oct 10. Proc Natl Acad Sci U S A. 2011. PMID: 21987786 Free PMC article.
-
Functions and regulation of the Polo-like kinase Cdc5 in the absence and presence of DNA damage.Curr Genet. 2018 Feb;64(1):87-96. doi: 10.1007/s00294-017-0727-2. Epub 2017 Aug 2. Curr Genet. 2018. PMID: 28770345 Free PMC article. Review.
Cited by
-
Saccharomyces cerevisiae Dbf4 has unique fold necessary for interaction with Rad53 kinase.J Biol Chem. 2012 Jan 20;287(4):2378-87. doi: 10.1074/jbc.M111.233973. Epub 2011 Nov 29. J Biol Chem. 2012. PMID: 22130670 Free PMC article.
-
The Polo kinase Cdc5 is regulated at multiple levels in the adaptation response to telomere dysfunction.Genetics. 2023 Jan 12;223(1):iyac171. doi: 10.1093/genetics/iyac171. Genetics. 2023. PMID: 36342193 Free PMC article.
-
Binding of Drosophila Polo kinase to its regulator Matrimony is noncanonical and involves two separate functional domains.Proc Natl Acad Sci U S A. 2013 Mar 26;110(13):E1222-31. doi: 10.1073/pnas.1301690110. Epub 2013 Mar 11. Proc Natl Acad Sci U S A. 2013. PMID: 23479640 Free PMC article.
-
An acidic loop in the forkhead-associated domain of the yeast meiosis-specific kinase Mek1 interacts with a specific motif in a subset of Mek1 substrates.Genetics. 2024 Sep 4;228(1):iyae106. doi: 10.1093/genetics/iyae106. Genetics. 2024. PMID: 38979911 Free PMC article.
-
Dbf4: the whole is greater than the sum of its parts.Cell Cycle. 2013 Apr 15;12(8):1180-8. doi: 10.4161/cc.24416. Epub 2013 Apr 2. Cell Cycle. 2013. PMID: 23549174 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
