

Functional dissection of LIS1 and NDEL1 towards understanding the molecular mechanisms of cytoplasmic dynein regulation
- PMID: 21036906
- PMCID: PMC3023492
- DOI: 10.1074/jbc.M110.169847
Functional dissection of LIS1 and NDEL1 towards understanding the molecular mechanisms of cytoplasmic dynein regulation
Abstract
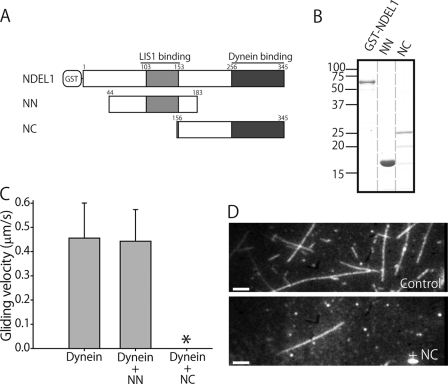
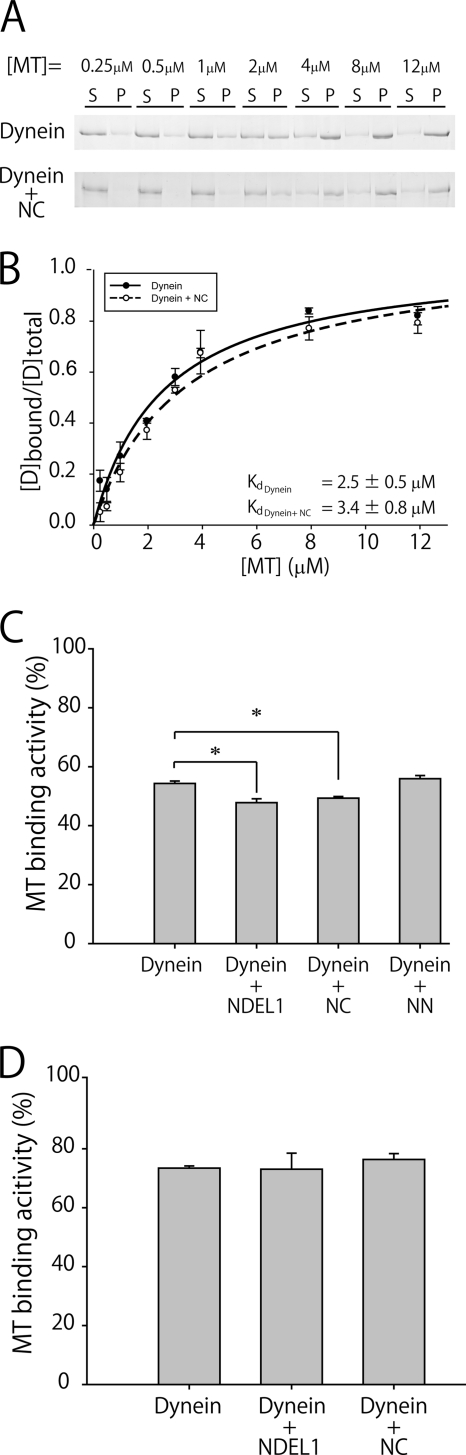
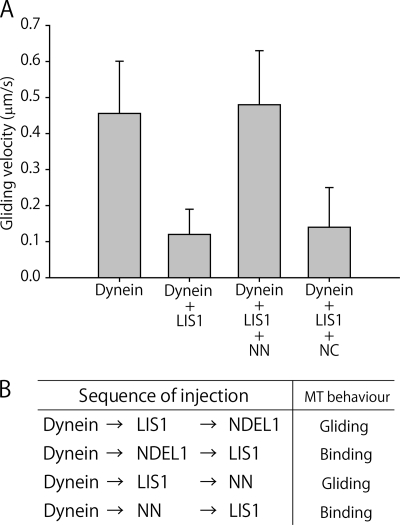
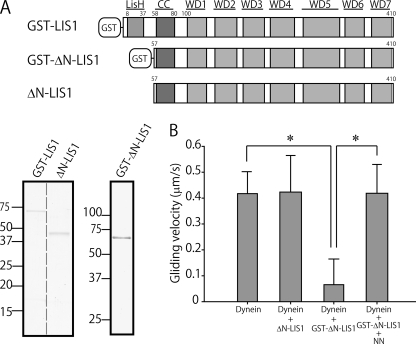
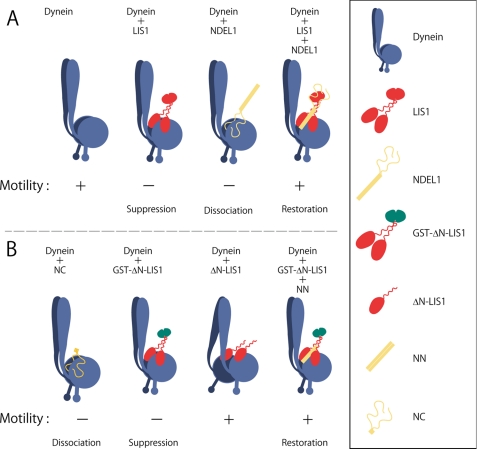
LIS1 and NDEL1 are known to be essential for the activity of cytoplasmic dynein in living cells. We previously reported that LIS1 and NDEL1 directly regulated the motility of cytoplasmic dynein in an in vitro motility assay. LIS1 suppressed dynein motility and inhibited the translocation of microtubules (MTs), while NDEL1 dissociated dynein from MTs and restored dynein motility following suppression by LIS1. However, the molecular mechanisms and detailed interactions of dynein, LIS1, and NDEL1 remain unknown. In this study, we dissected the regulatory effects of LIS1 and NDEL1 on dynein motility using full-length or truncated recombinant fragments of LIS1 or NDEL1. The C-terminal fragment of NDEL1 dissociated dynein from MTs, whereas its N-terminal fragment restored dynein motility following suppression by LIS1, demonstrating that the two functions of NDEL1 localize to different parts of the NDEL1 molecule, and that restoration from LIS1 suppression is caused by the binding of NDEL1 to LIS1, rather than to dynein. The truncated monomeric form of LIS1 had little effect on dynein motility, but an artificial dimer of truncated LIS1 suppressed dynein motility, which was restored by the N-terminal fragment of NDEL1. This suggests that LIS1 dimerization is essential for its regulatory function. These results shed light on the molecular interactions between dynein, LIS1, and NDEL1, and the mechanisms of cytoplasmic dynein regulation.
Figures
References
-
- Vallee R. B., Williams J. C., Varma D., Barnhart L. E. (2004) J. Neurobiol. 58, 189–200 - PubMed
-
- Schroer T. A. (2004) Annu. Rev. Cell Dev. Biol. 20, 759–779 - PubMed
-
- Reiner O., Carrozzo R., Shen Y., Wehnert M., Faustinella F., Dobyns W. B., Caskey C. T., Ledbetter D. H. (1993) Nature 364, 717–721 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
