

High-performance spider webs: integrating biomechanics, ecology and behaviour
- PMID: 21036911
- PMCID: PMC3061126
- DOI: 10.1098/rsif.2010.0454
High-performance spider webs: integrating biomechanics, ecology and behaviour
Abstract
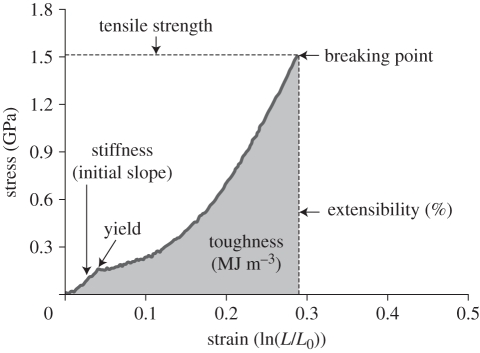
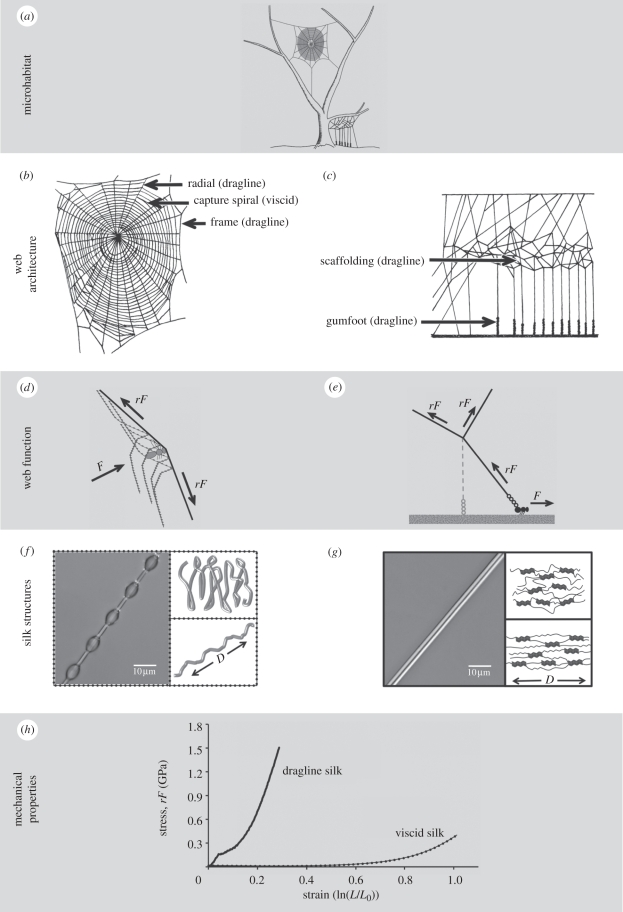
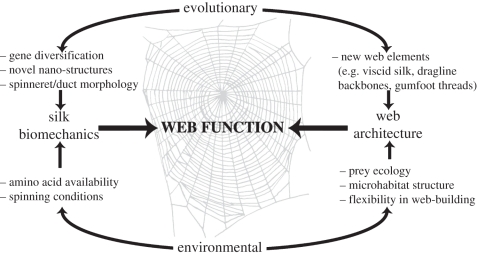
Spider silks exhibit remarkable properties, surpassing most natural and synthetic materials in both strength and toughness. Orb-web spider dragline silk is the focus of intense research by material scientists attempting to mimic these naturally produced fibres. However, biomechanical research on spider silks is often removed from the context of web ecology and spider foraging behaviour. Similarly, evolutionary and ecological research on spiders rarely considers the significance of silk properties. Here, we highlight the critical need to integrate biomechanical and ecological perspectives on spider silks to generate a better understanding of (i) how silk biomechanics and web architectures interacted to influence spider web evolution along different structural pathways, and (ii) how silks function in an ecological context, which may identify novel silk applications. An integrative, mechanistic approach to understanding silk and web function, as well as the selective pressures driving their evolution, will help uncover the potential impacts of environmental change and species invasions (of both spiders and prey) on spider success. Integrating these fields will also allow us to take advantage of the remarkable properties of spider silks, expanding the range of possible silk applications from single threads to two- and three-dimensional thread networks.
Figures
References
-
- Hansell M. H. 2005. Animal architecture. 1st edn. New York, NY: Oxford University Press
-
- Madin J. S., Connolly S. R. 2006. Ecological consequences of major hydrodynamic disturbances on coral reefs. Nature 444, 477–48010.1038/nature05328 (doi:10.1038/nature05328) - DOI - DOI - PubMed
-
- Palmer A. R. 1979. Fish predation and evolution of gastropod shell sculpture: experimental and geographic evidence. Evolution 33, 697–71310.2307/2407792 (doi:10.2307/2407792) - DOI - DOI - PubMed
-
- Craig C. L. 1987. The ecological and evolutionary interdependence between web architecture and web silk spun by orb web weaving spiders. Biol. J. Linn. Soc. 30, 135–16210.1111/j.1095-8312.1987.tb00294.x (doi:10.1111/j.1095-8312.1987.tb00294.x) - DOI - DOI
-
- Gosline J. M., Guerette P. A., Ortlepp C. S., Savage K. N. 1999. The mechanical design of spider silks: from fibroin sequence to mechanical function. J. Exp. Biol. 202, 3295–3303 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
