

Discovery of non-ETS gene fusions in human prostate cancer using next-generation RNA sequencing
- PMID: 21036922
- PMCID: PMC3012926
- DOI: 10.1101/gr.110684.110
Discovery of non-ETS gene fusions in human prostate cancer using next-generation RNA sequencing
Abstract
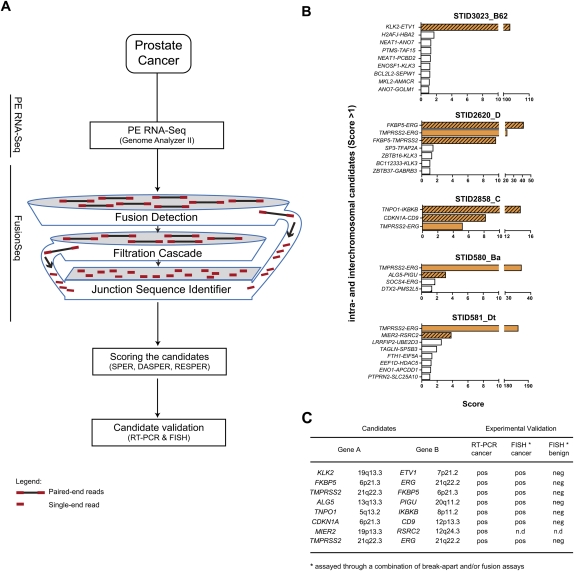
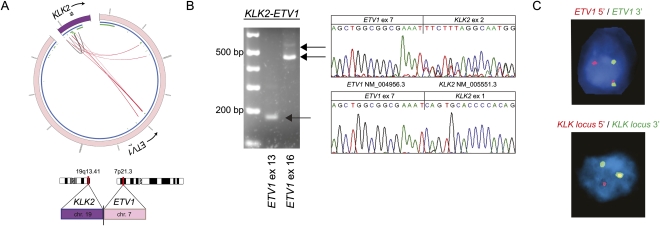
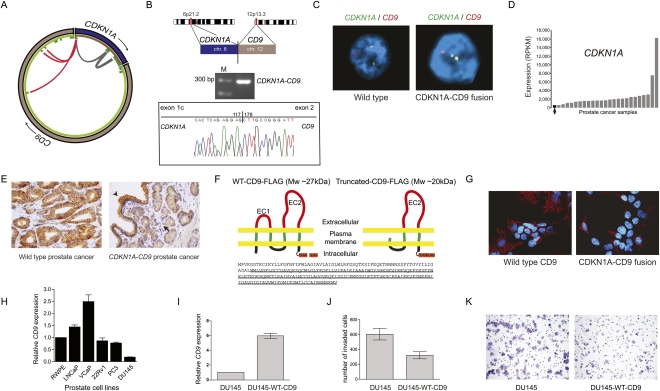
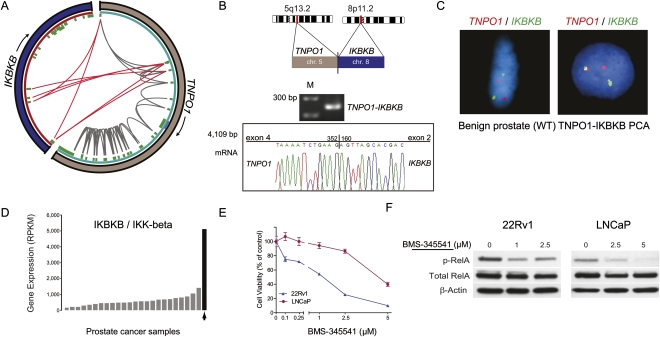
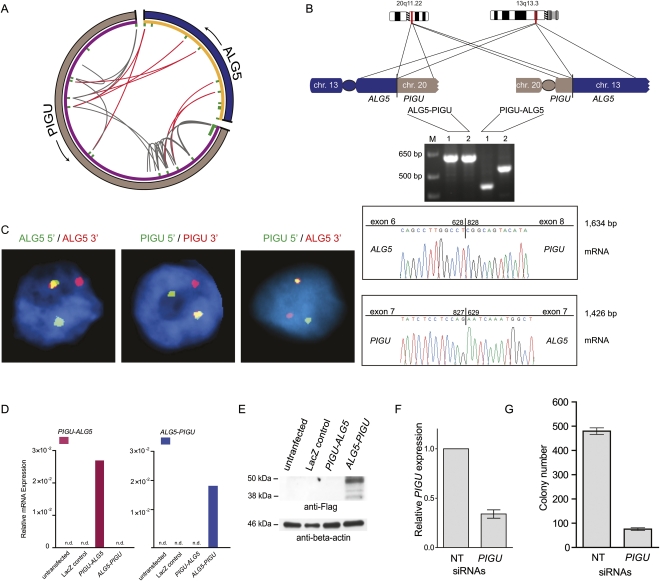
Half of prostate cancers harbor gene fusions between TMPRSS2 and members of the ETS transcription factor family. To date, little is known about the presence of non-ETS fusion events in prostate cancer. We used next-generation transcriptome sequencing (RNA-seq) in order to explore the whole transcriptome of 25 human prostate cancer samples for the presence of chimeric fusion transcripts. We generated more than 1 billion sequence reads and used a novel computational approach (FusionSeq) in order to identify novel gene fusion candidates with high confidence. In total, we discovered and characterized seven new cancer-specific gene fusions, two involving the ETS genes ETV1 and ERG, and four involving non-ETS genes such as CDKN1A (p21), CD9, and IKBKB (IKK-beta), genes known to exhibit key biological roles in cellular homeostasis or assumed to be critical in tumorigenesis of other tumor entities, as well as the oncogene PIGU and the tumor suppressor gene RSRC2. The novel gene fusions are found to be of low frequency, but, interestingly, the non-ETS fusions were all present in prostate cancer harboring the TMPRSS2-ERG gene fusion. Future work will focus on determining if the ETS rearrangements in prostate cancer are associated or directly predispose to a rearrangement-prone phenotype.
Figures
References
-
- Bergsagel PL, Kuehl WM 2001. Chromosome translocations in multiple myeloma. Oncogene 20: 5611–5622 - PubMed
-
- Burke JR, Pattoli MA, Gregor KR, Brassil PJ, MacMaster JF, McIntyre KW, Yang X, Iotzova VS, Clarke W, Strnad J, et al. 2003. BMS-345541 is a highly selective inhibitor of IκB kinase that binds at an allosteric site of the enzyme and blocks NF-κB-dependent transcription in mice. J Biol Chem 278: 1450–1456 - PubMed
-
- Chariot A 2009. The NF-κB-independent functions of IKK subunits in immunity and cancer. Trends Cell Biol 19: 404–413 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases