

Hematopoietic differentiation of induced pluripotent stem cells from patients with mucopolysaccharidosis type I (Hurler syndrome)
- PMID: 21037085
- PMCID: PMC3035077
- DOI: 10.1182/blood-2010-05-287607
Hematopoietic differentiation of induced pluripotent stem cells from patients with mucopolysaccharidosis type I (Hurler syndrome)
Abstract
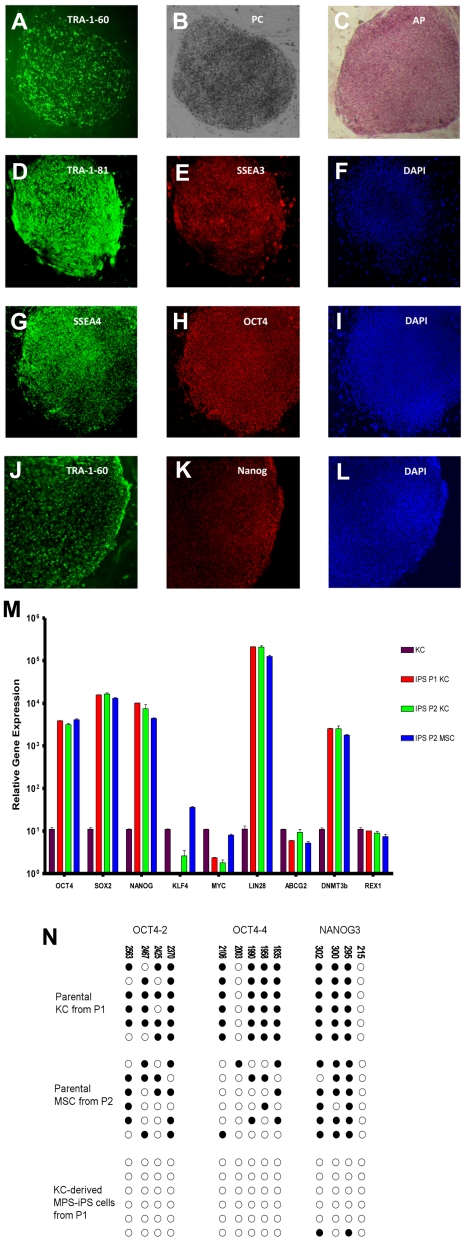
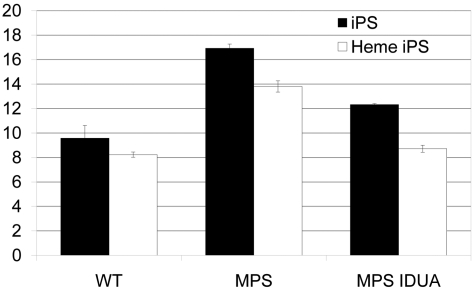
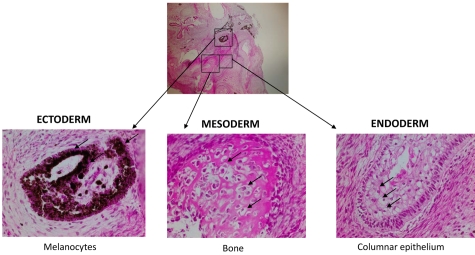
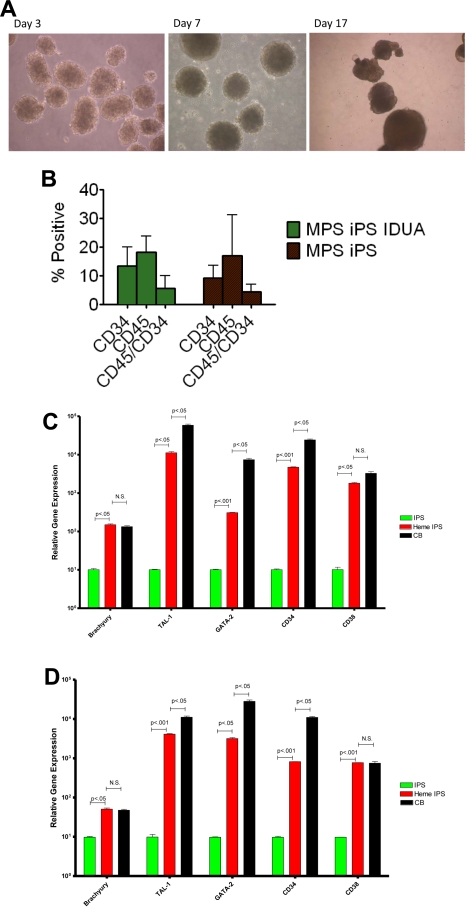
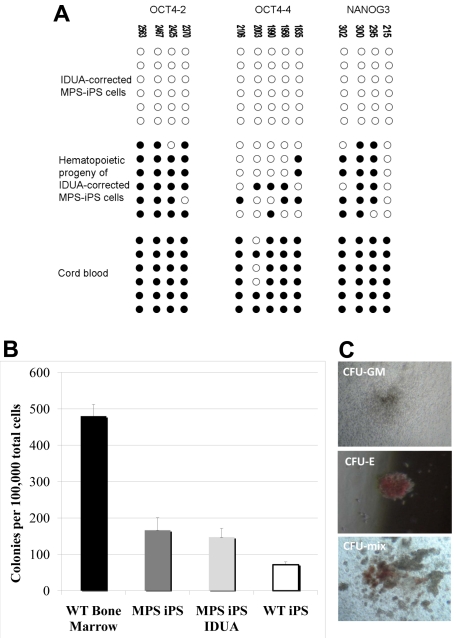
Mucopolysaccharidosis type I (MPS IH; Hurler syndrome) is a congenital deficiency of α-L-iduronidase, leading to lysosomal storage of glycosaminoglycans that is ultimately fatal following an insidious onset after birth. Hematopoietic cell transplantation (HCT) is a life-saving measure in MPS IH. However, because a suitable hematopoietic donor is not found for everyone, because HCT is associated with significant morbidity and mortality, and because there is no known benefit of immune reaction between the host and the donor cells in MPS IH, gene-corrected autologous stem cells may be the ideal graft for HCT. Thus, we generated induced pluripotent stem cells from 2 patients with MPS IH (MPS-iPS cells). We found that α-L-iduronidase was not required for stem cell renewal, and that MPS-iPS cells showed lysosomal storage characteristic of MPS IH and could be differentiated to both hematopoietic and nonhematopoietic cells. The specific epigenetic profile associated with de-differentiation of MPS IH fibroblasts into MPS-iPS cells was maintained when MPS-iPS cells are gene-corrected with virally delivered α-L-iduronidase. These data underscore the potential of MPS-iPS cells to generate autologous hematopoietic grafts devoid of immunologic complications of allogeneic transplantation, as well as generating nonhematopoietic cells with the potential to treat anatomical sites not fully corrected with HCT.
Figures
References
-
- Fratantoni JC, Hall CW, Neufeld EF. Hurler and Hunter syndromes: mutual correction of the defect in cultured fibroblasts. Science. 1968;162(853):570–572. - PubMed
-
- Hobbs JR, Hugh-Jones K, Barrett AJ, et al. Reversal of clinical features of Hurler's disease and biochemical improvement after treatment by bone-marrow transplantation. Lancet. 1981;2(8249):709–712. - PubMed
-
- Orchard PJ, Blazar BR, Wagner J, Charnas L, Krivit W, Tolar J. Hematopoietic cell therapy for metabolic disease. J Pediatr. 2007;151(4):340–346. - PubMed
-
- Kakkis ED, Muenzer J, Tiller GE, et al. Enzyme-replacement therapy in mucopolysaccharidosis I. N Engl J Med. 2001;344(3):182–188. - PubMed
-
- Krivit W, Sung JH, Shapiro EG, Lockman LA. Microglia: the effector cell for reconstitution of the central nervous system following bone marrow transplantation for lysosomal and peroxisomal storage diseases. Cell Transplant. 1995;4(4):385–392. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
