

LFA-1 and Mac-1 define characteristically different intralumenal crawling and emigration patterns for monocytes and neutrophils in situ
- PMID: 21037096
- PMCID: PMC3004223
- DOI: 10.4049/jimmunol.1001638
LFA-1 and Mac-1 define characteristically different intralumenal crawling and emigration patterns for monocytes and neutrophils in situ
Abstract
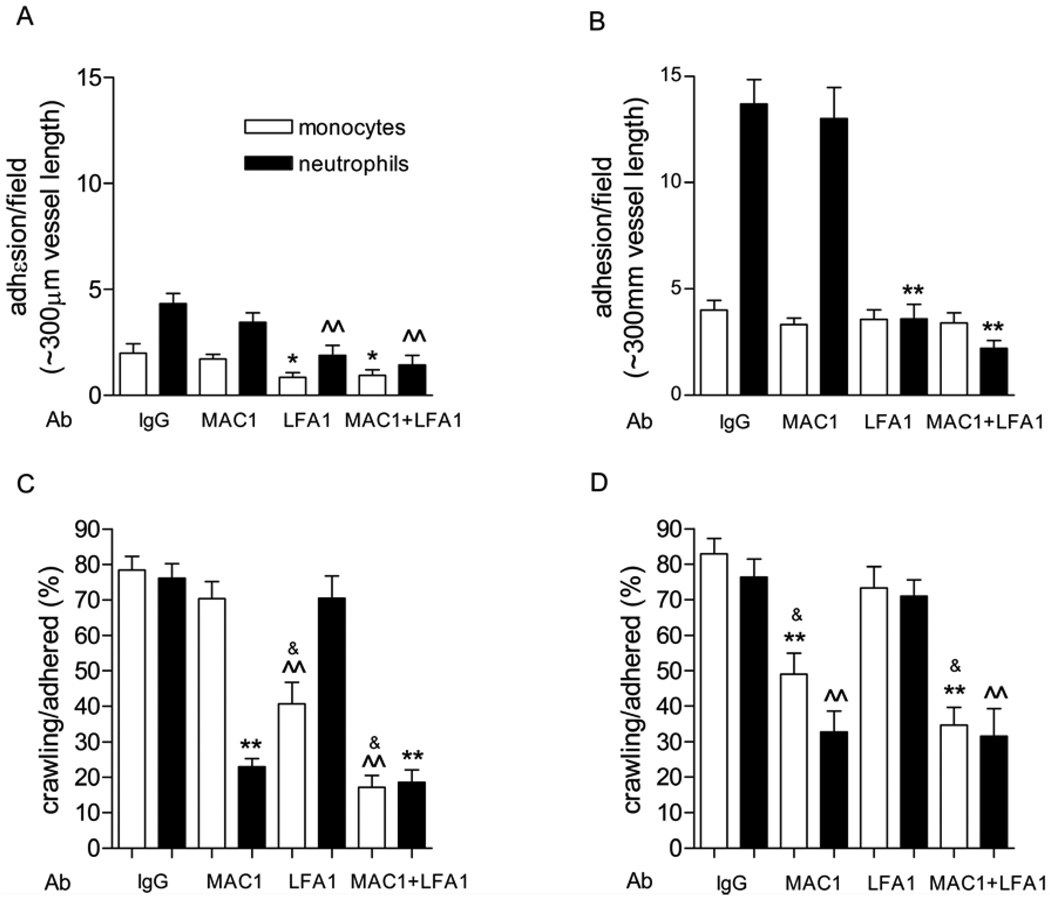
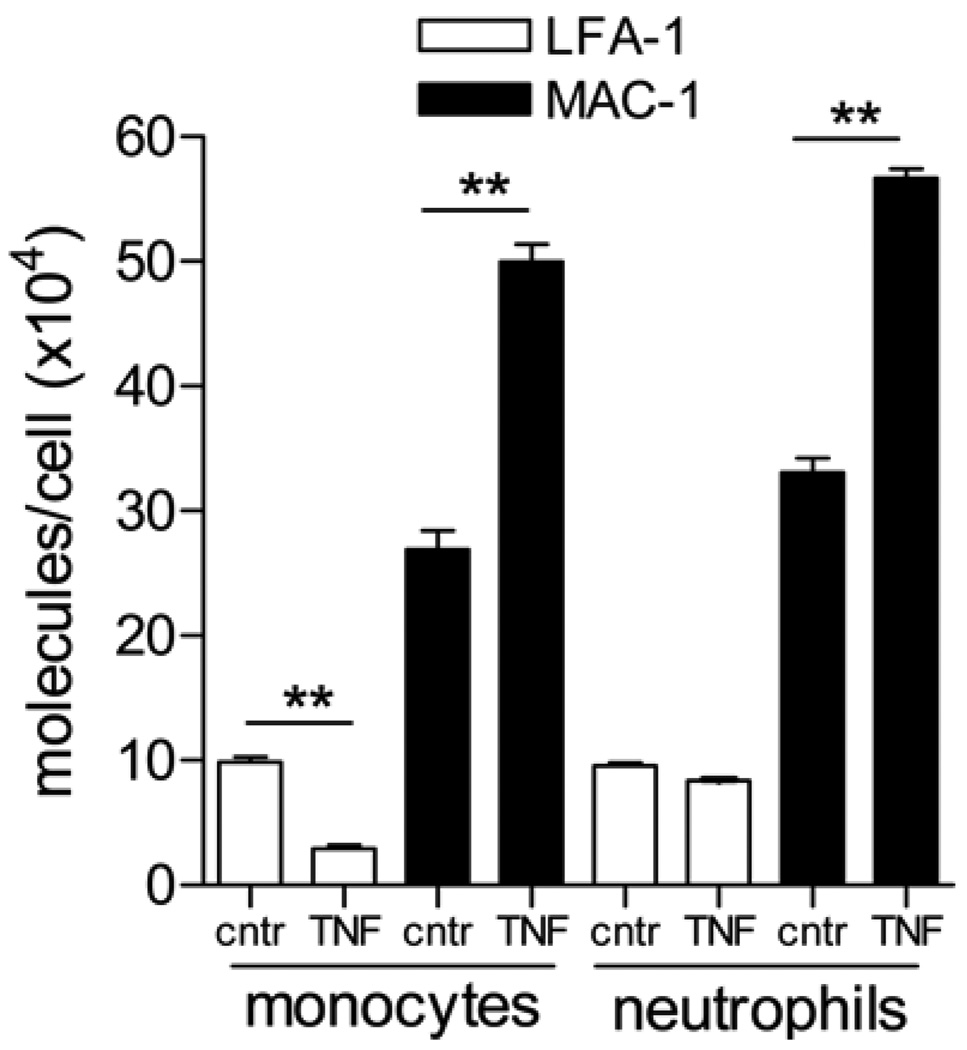
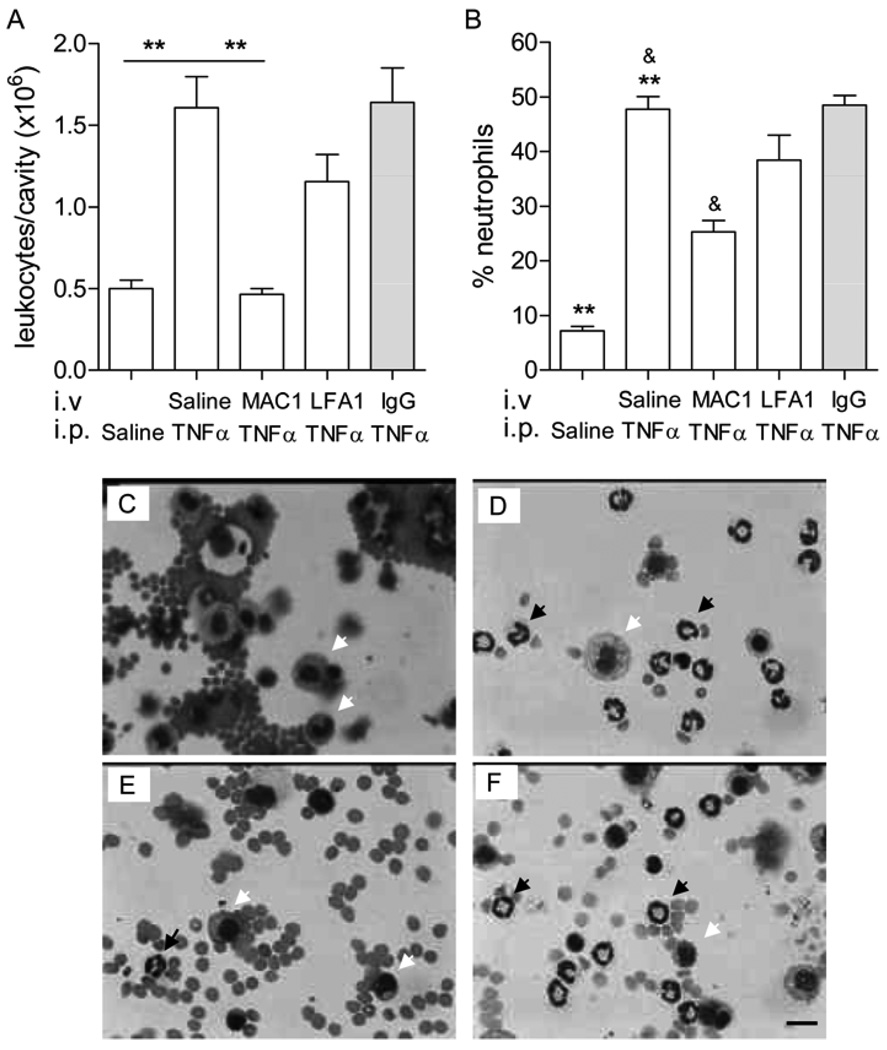
To exit blood vessels, most (∼80%) of the lumenally adhered monocytes and neutrophils crawl toward locations that support transmigration. Using intravital confocal microscopy of anesthetized mouse cremaster muscle, we separately examined the crawling and emigration patterns of monocytes and neutrophils in blood-perfused unstimulated or TNF-α-activated venules. Most of the interacting cells in microvessels are neutrophils; however, in unstimulated venules, a greater percentage of the total monocyte population is adherent compared with neutrophils (58.2 ± 6.1% versus 13.6 ± 0.9%, adhered/total interacting), and they crawl for significantly longer distances (147.3 ± 13.4 versus 61.8 ± 5.4 μm). Intriguingly, after TNF-α activation, monocytes crawled for significantly shorter distances (67.4 ± 9.6 μm), resembling neutrophil crawling. Using function-blocking Abs, we show that these different crawling patterns were due to CD11a/CD18 (LFA-1)- versus CD11b/CD18 (Mac-1)-mediated crawling. Blockade of either Mac-1 or LFA-1 revealed that both LFA-1 and Mac-1 contribute to monocyte crawling; however, the LFA-1-dependent crawling in unstimulated venules becomes Mac-1 dependent upon inflammation, likely due to increased expression of Mac-1. Mac-1 alone was responsible for neutrophil crawling in both unstimulated and TNF-α-activated venules. Consistent with the role of Mac-1 in crawling, Mac-1 block (compared with LFA-1) was also significantly more efficient in blocking TNF-α-induced extravasation of both monocytes and neutrophils in cremaster tissue and the peritoneal cavity. Thus, mechanisms underlying leukocyte crawling are important in regulating the inflammatory responses by regulating the numbers of leukocytes that transmigrate.
Figures




References
-
- Smith JA. Neutrophils, host defense, and inflammation: a double-edged sword. J Leukoc Biol. 1994;56:672–686. - PubMed
-
- Muller WA, Randolph GJ. Migration of leukocytes across endothelium and beyond: molecules involved in the transmigration and fate of monocytes. J Leukoc Biol. 1999;66:698–704. - PubMed
-
- Kansas GS. Selectins and their ligands: current concepts and controversies. Blood. 1996;88:3259–3287. - PubMed
-
- Ley K, Allietta M, Bullard DC, Morgan S. Importance of E-selectin for firm leukocyte adhesion in vivo. Circ Res. 1998;83:287–294. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
