

doi: 10.4161/rna.7.5.13097.
Epub 2010 Sep 1.
Regulation of virulence gene expression in Streptococcus pyogenes: determinants of differential mRNA decay
Affiliations
- PMID: 21037420
- PMCID: PMC3073253
- DOI: 10.4161/rna.7.5.13097
Item in Clipboard
Regulation of virulence gene expression in Streptococcus pyogenes: determinants of differential mRNA decay
RNA Biol.
2010 Sep-Oct.
Abstract
Differential mRNA stability is an important mechanism for regulation of virulence factors in Streptococcus pyogenes (group A streptococcus, GAS), a serious and prevalent human pathogen. We have described 2 Classes of mRNA in GAS that are distinguishable by 1) stability in the stationary phase of growth, 2) kinetics of decay in exponential phase, and 3) effect of depletion of RNases J1 and J2 and polynucleotide phosphorylase (PNPase) on decay in exponential phase. We discuss features of the structure of an mRNA that appear to be important for determining the Class to which it belongs and present a model to explain differential mRNA decay.
Figures
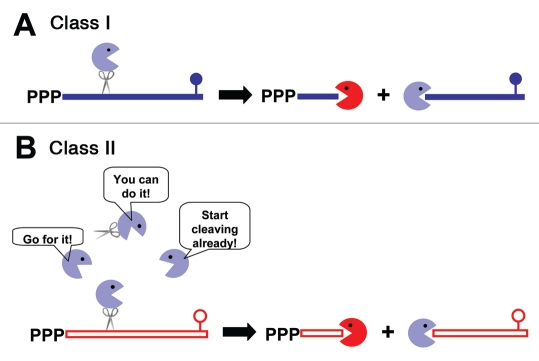
Models for exponential phase mRNA decay in GAS. (A) The 5′ end of Class I mRNA (shown in blue) makes it a good substrate for RNase J1 and/or RNase J2 (blue pacman). Cleavage is most likely initiated by endonucleolytic activity of these RNases, which produces two type of products. One has an accessible 3′-end and is subject to digestion by 3′-to-5′ exonucleases (red pacman) and the other has a 5′-monophosphate which makes it a good substrate for the 5′-to-3′ exonucleolytic activity of the J RNases. (B) Class II mRNA (shown in red) is less sensitive to the J RNases, so cleavage of these mRNAs does not begin until Class I messages are depleted, freeing RNase J1 and J2 to bind to Class II mRNAs. Cleavage of Class II mRNAs is initiated endonucleolytically by the J RNases, releasing products similar to those produced from Class I mRNAs. These products are degraded 5′-to-3′ by RNase J1 and J2 and 3′-to-5′ by the 3′-to-5′ exonucleases, predominantly PNPase.
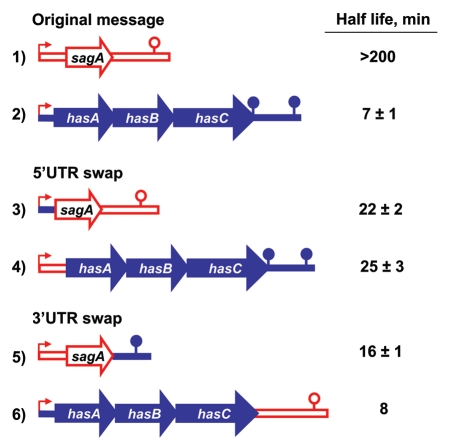
Decay rates of chimeric messages. The sag transcript is represented in red and the has transcript in blue. All transcripts are produced from Psag. The 5′-UTR begins with the transcription start, includes the ribosome binding site and ends just upstream of the translation initiation codon. The next segment starts with the translation initiation codon (AUG for sag and GUG for has), ends with the translation stop codon and comprises sagA for sag or hasABC for has. The 3′-UTR starts immediately after the translation stop codon and includes the predicted transcription pause site for sag or transcription terminator for has (lollipop). The messages were assembled using overlapping PCR and cloned under the Psag promoter (bent arrow) into plasmid pJRS9508 to generate plasmids: (1) pJRS2187; (2) pJRS1293, (3) pEU7745; (4) pEU7748; (5) pEU7753; (6) pEU7754. The chimeric constructs were expressed in the GAS strain JRS1288 (Δcov Δsag MGAS315) as described. GAS was grown to 1 hour into stationary phase in Todd Hewitt broth with pH adjusted to 7.5 with 100 mM HEPES, transcription was stopped by addition of rifampicin to the culture and RNA was isolated from cells collected at four to six consequent time points after rifampicin addition as described. The amounts for sag and has messages in each RNA sample were determined by Q-RT-PCR (four technical replicates for each sample) with primers designed to detect the middle part of the translated portion for sagA or hasA mRNA. RNA isolated from at least two independent cultures was used for Q-RT-PCR for constructs 1 to 5 and from a single culture for construct 6. Decay rates were confirmed using northern blot analysis, for constructs in lines 1, 2 and 4.
Similar articles
-
The ribonucleases J1 and J2 are essential for growth and have independent roles in mRNA decay in Streptococcus pyogenes.Mol Microbiol. 2010 Feb;75(3):731-43. doi: 10.1111/j.1365-2958.2009.07012.x. Epub 2009 Dec 16. Mol Microbiol. 2010. PMID: 20025665
-
Role of mRNA stability in growth phase regulation of gene expression in the group A streptococcus.J Bacteriol. 2007 Mar;189(5):1866-73. doi: 10.1128/JB.01658-06. Epub 2006 Dec 22. J Bacteriol. 2007. PMID: 17189377 Free PMC article.
-
Endopeptidase PepO Regulates the SpeB Cysteine Protease and Is Essential for the Virulence of Invasive M1T1 Streptococcus pyogenes.J Bacteriol. 2018 Mar 26;200(8):e00654-17. doi: 10.1128/JB.00654-17. Print 2018 Apr 15. J Bacteriol. 2018. PMID: 29378883 Free PMC article.
-
Virulence factor regulation and regulatory networks in Streptococcus pyogenes and their impact on pathogen-host interactions.Trends Microbiol. 2003 May;11(5):224-32. doi: 10.1016/s0966-842x(03)00098-2. Trends Microbiol. 2003. PMID: 12781526 Review.
-
Dissecting Streptococcus pyogenes interaction with human.Arch Microbiol. 2020 Oct;202(8):2023-2032. doi: 10.1007/s00203-020-01932-w. Epub 2020 Jun 5. Arch Microbiol. 2020. PMID: 32504132 Review.
Cited by
-
A newly identified duplex RNA unwinding activity of archaeal RNase J depends on processive exoribonucleolysis coupled steric occlusion by its structural archaeal loops.RNA Biol. 2020 Oct;17(10):1480-1491. doi: 10.1080/15476286.2020.1777379. Epub 2020 Jun 18. RNA Biol. 2020. PMID: 32552320 Free PMC article.
-
covR Mediated Antibiofilm Activity of 3-Furancarboxaldehyde Increases the Virulence of Group A Streptococcus.PLoS One. 2015 May 15;10(5):e0127210. doi: 10.1371/journal.pone.0127210. eCollection 2015. PLoS One. 2015. PMID: 25978065 Free PMC article.
-
Streptococcus pyogenes biofilms-formation, biology, and clinical relevance.Front Cell Infect Microbiol. 2015 Feb 11;5:15. doi: 10.3389/fcimb.2015.00015. eCollection 2015. Front Cell Infect Microbiol. 2015. PMID: 25717441 Free PMC article. Review.
-
When ribonucleases come into play in pathogens: a survey of gram-positive bacteria.Int J Microbiol. 2012;2012:592196. doi: 10.1155/2012/592196. Epub 2012 Mar 13. Int J Microbiol. 2012. PMID: 22550495 Free PMC article.
-
Mab_3083c Is a Homologue of RNase J and Plays a Role in Colony Morphotype, Aggregation, and Sliding Motility of Mycobacterium abscessus.Microorganisms. 2021 Mar 25;9(4):676. doi: 10.3390/microorganisms9040676. Microorganisms. 2021. PMID: 33805851 Free PMC article.
References
-
- Cunningham MW. Pathogenesis of group A streptococcal infections and their sequelae. Advances in experimental medicine and biology. 2008;609:2942. - PubMed
-
- Tart AH, Walker MJ, Musser JM. New understanding of the group A Streptococcus pathogenesis cycle. Trends in microbiology. 2007;15:318–325. - PubMed
-
- Carapetis JR, Steer AC, Mulholland EK, Weber M. The global burden of group A streptococcal diseases. The Lancet infectious diseases. 2005;5:685–694. - PubMed
-
- Martin JM, Green M. Group A streptococcus. Seminars in pediatric infectious diseases. 2006;17:140–148. - PubMed
-
- Kaplan EL, Johnson DR. Unexplained reduced microbiological efficacy of intramuscular benzathine penicillin G and of oral penicillin V in eradication of group a streptococci from children with acute pharyngitis. Pediatrics. 2001;108:1180–1186. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources