

Maltose-neopentyl glycol (MNG) amphiphiles for solubilization, stabilization and crystallization of membrane proteins
- PMID: 21037590
- PMCID: PMC3063152
- DOI: 10.1038/nmeth.1526
Maltose-neopentyl glycol (MNG) amphiphiles for solubilization, stabilization and crystallization of membrane proteins
Abstract
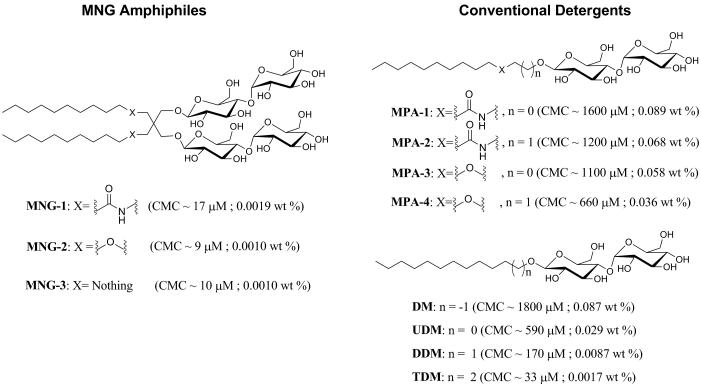
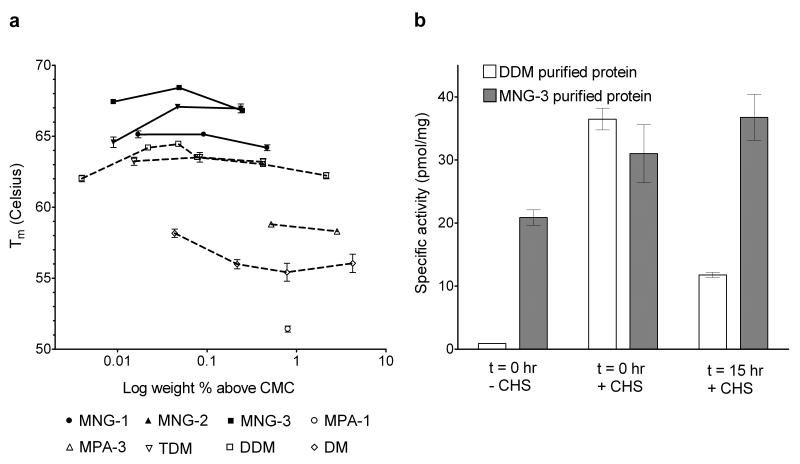
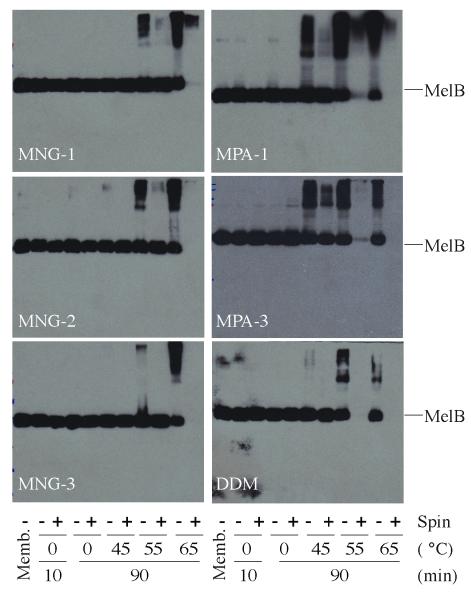
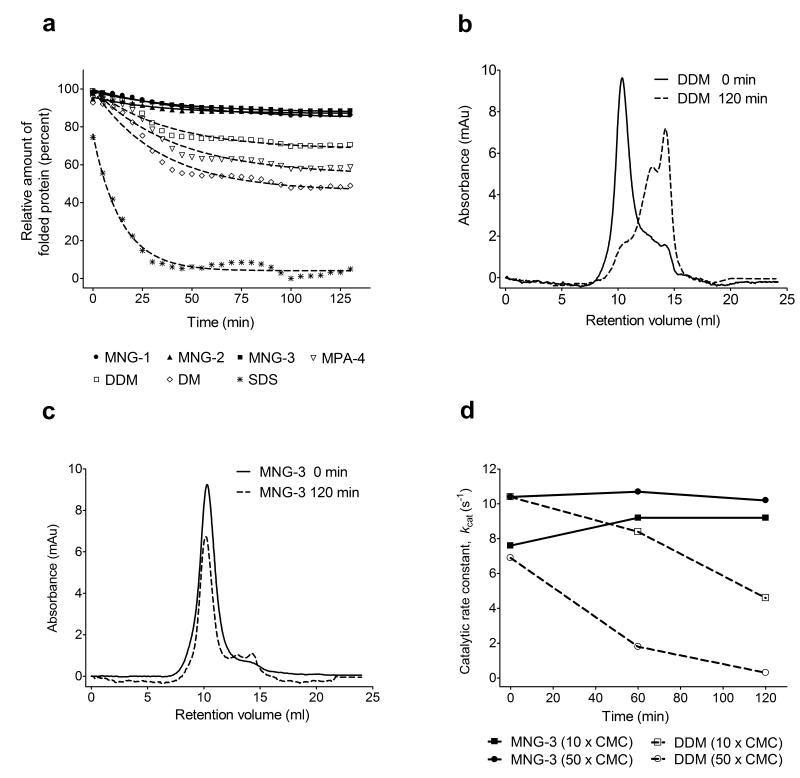
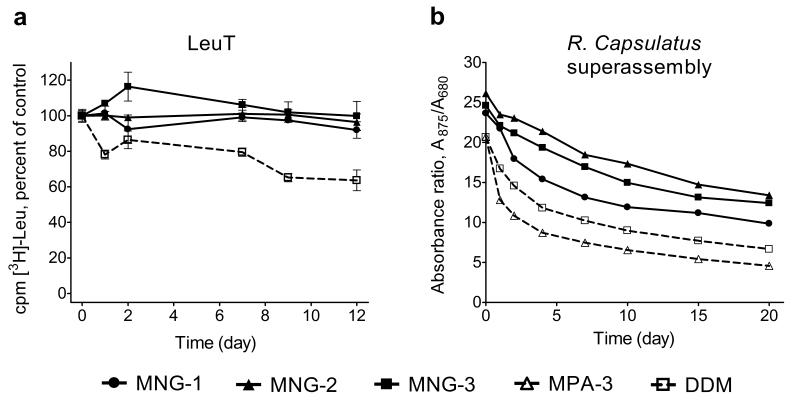
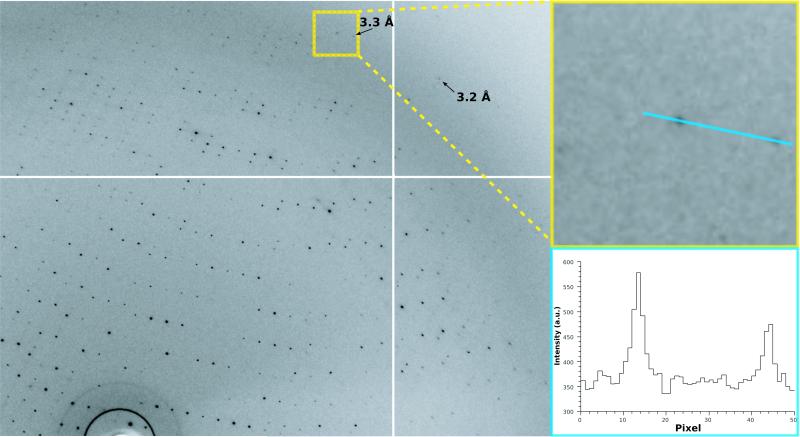
The understanding of integral membrane protein (IMP) structure and function is hampered by the difficulty of handling these proteins. Aqueous solubilization, necessary for many types of biophysical analysis, generally requires a detergent to shield the large lipophilic surfaces of native IMPs. Many proteins remain difficult to study owing to a lack of suitable detergents. We introduce a class of amphiphiles, each built around a central quaternary carbon atom derived from neopentyl glycol, with hydrophilic groups derived from maltose. Representatives of this maltose-neopentyl glycol (MNG) amphiphile family show favorable behavior relative to conventional detergents, as manifested in multiple membrane protein systems, leading to enhanced structural stability and successful crystallization. MNG amphiphiles are promising tools for membrane protein science because of the ease with which they may be prepared and the facility with which their structures may be varied.
Figures
References
-
- Lacapere JJ, Pebay-Peyroula E, Neumann JM, Etchebest C. Determining membrane protein structures: still a challenge! Trends Biochem. Sci. 2007;32:259–270. - PubMed
-
- Privé GG. Detergents for the stabilization and crystallization of membrane proteins. Methods. 2007;41:388–397. - PubMed
-
- Schafmeister CE, Meircke LJW, Stroud RM. Structure at 2.5 Å of a designed peptide that maintains solubility of membrane proteins. Science. 1993;262:734–738. - PubMed
-
- McGregor C-L, et al. Lipopeptide detergents designed for the structural study of membrane protein. Nat. Biotech. 2003;21:171–176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R21 HL087895/HL/NHLBI NIH HHS/United States
- U54 GM-074901/GM/NIGMS NIH HHS/United States
- GM083118/GM/NIGMS NIH HHS/United States
- R01 GM050853/GM/NIGMS NIH HHS/United States
- R01 GM083118/GM/NIGMS NIH HHS/United States
- P01 GM075913/GM/NIGMS NIH HHS/United States
- R01 NS028471/NS/NINDS NIH HHS/United States
- U54 GM074901/GM/NIGMS NIH HHS/United States
- P01 NS28471/NS/NINDS NIH HHS/United States
- P01 GM75913/GM/NIGMS NIH HHS/United States
- R21HL087895/HL/NHLBI NIH HHS/United States
- R37 NS028471/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
