

Differences in T-helper polarizing capability between human monocyte-derived dendritic cells and monocyte-derived Langerhans'-like cells
- PMID: 21039466
- PMCID: PMC3050445
- DOI: 10.1111/j.1365-2567.2010.03356.x
Differences in T-helper polarizing capability between human monocyte-derived dendritic cells and monocyte-derived Langerhans'-like cells
Abstract
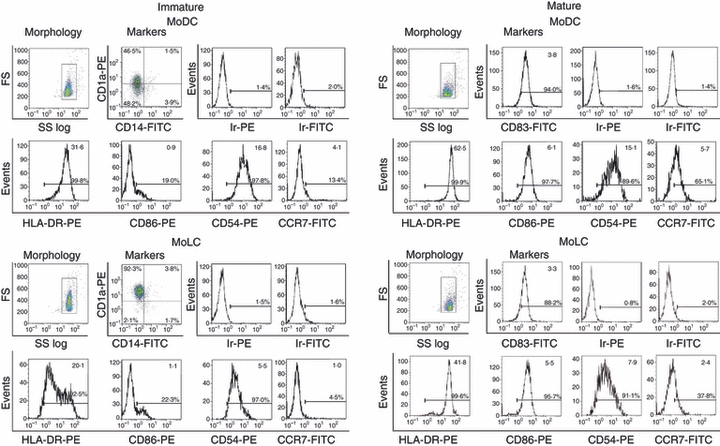
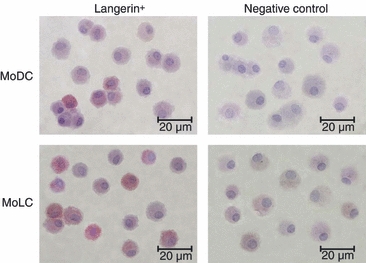
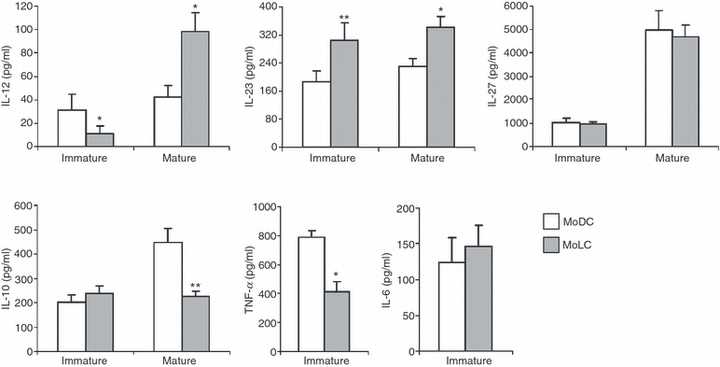
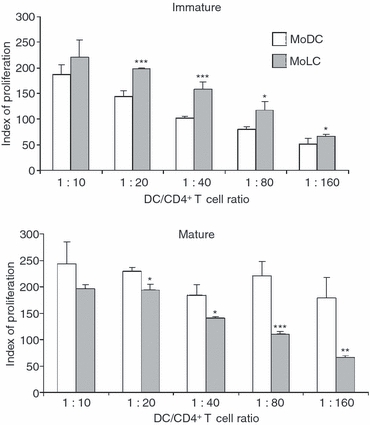
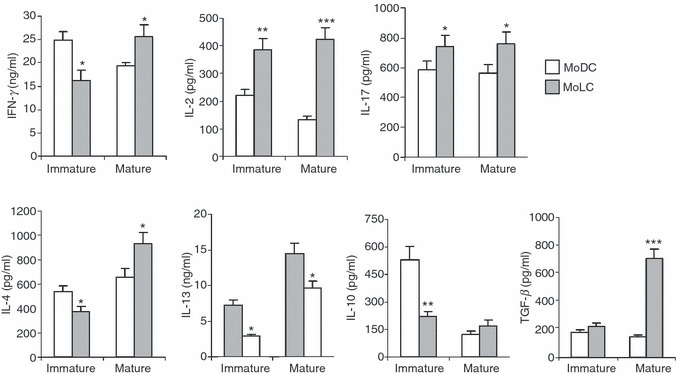
Langerhans' cells (LCs) represent a specific subset of dendritic cells (DCs) which are important for detecting and processing pathogens that penetrate the skin and epithelial barriers. The aim of our study was to explain what makes their in vitro counterparts - monocyte-derived Langerhans'-like cells (MoLCs) - unique compared with monocyte-derived dendritic cells (MoDCs). Immature MoDCs were generated by incubating peripheral blood monocytes with granulocyte-macrophage colony-stimulating factor (GM-CSF) and interleukin (IL)-4. The addition of transforming growth factor-β (TGF-β) to this cytokine cocktail resulted in the generation of MoLCs. MoLCs showed a lower expression of CD83, CD86, HLA-DR and CCR7 compared with MoDCs, regardless of their maturational status. Both immature and mature MoLCs secreted higher quantities of IL-23 compared with MoDCs and this finding correlated with a higher secretion of IL-17 in co-culture of MoLCs with allogeneic CD4(+) T cells. Mature MoLCs, which produced higher levels of IL-12 and lower levels of IL-10 compared with mature MoDCs, were more potent at inducing interferon-γ (IFN-γ) production by CD4(+) T cells in the co-culture system. In conclusion, the finding that mature MoLCs stimulate stronger T-helper 1 and T-helper 17 immune responses than mature MoDCs, makes them better candidates for use in the preparation of anti-tumour DC vaccines.
Figures
References
-
- Riedl E, Stockl J, Majdic O, Scheinecker C, Knapp W, Strobl H. Ligation of E-cadherin on in vitro-generated immature Langerhans-type dendritic cells inhibits their maturation. Blood. 2000;96:4276–84. - PubMed
-
- Banchereau J, Briere F, Caux C, et al. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
