

Wallerian degeneration of zebrafish trigeminal axons in the skin is required for regeneration and developmental pruning
- PMID: 21041367
- PMCID: PMC2976282
- DOI: 10.1242/dev.053611
Wallerian degeneration of zebrafish trigeminal axons in the skin is required for regeneration and developmental pruning
Abstract
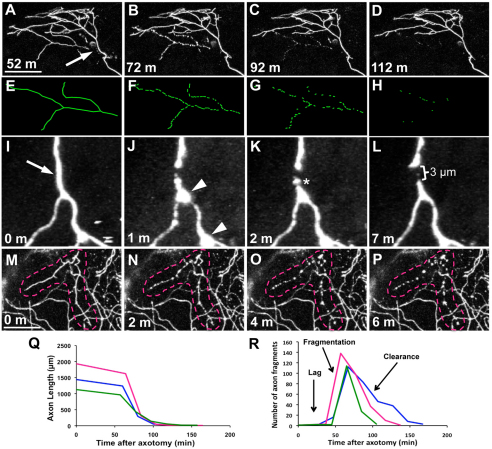
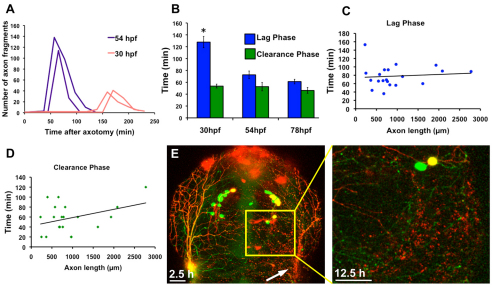
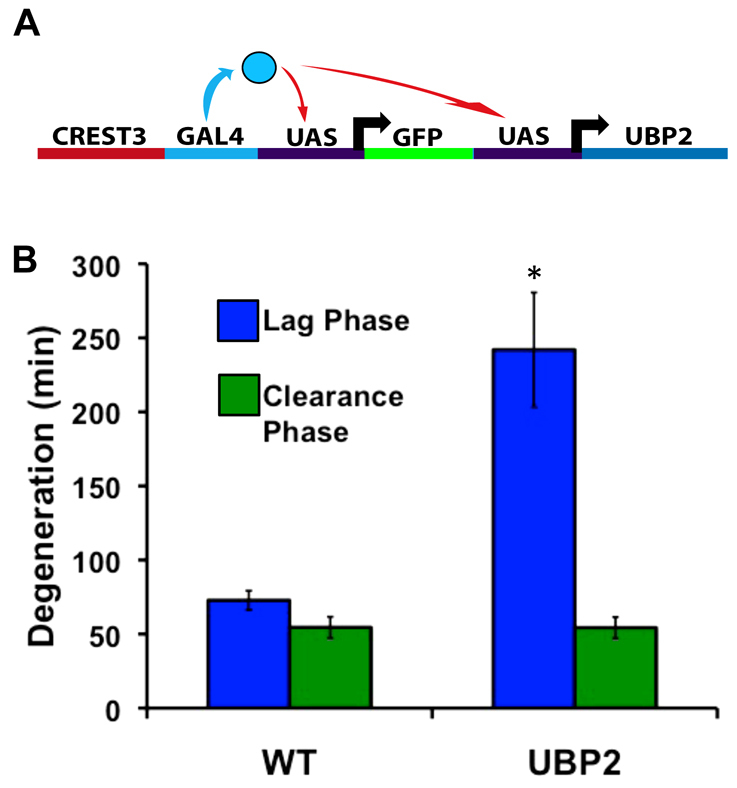
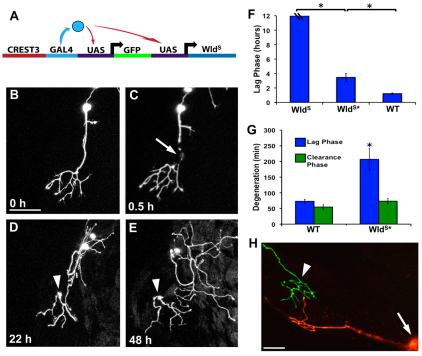
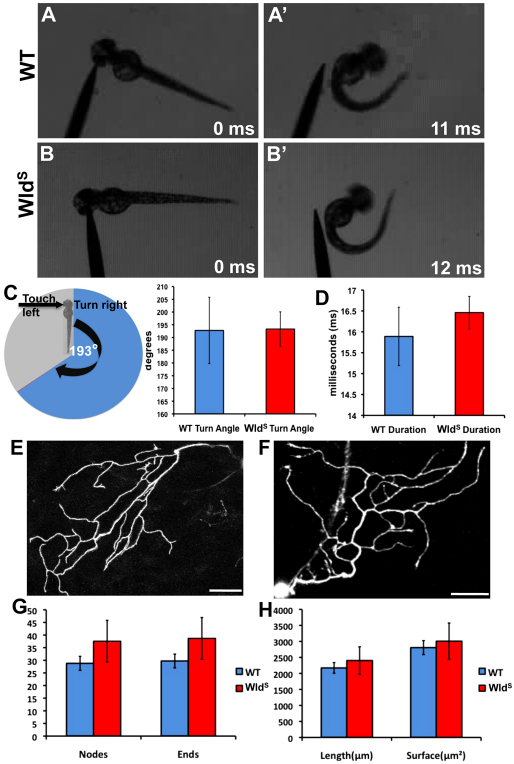
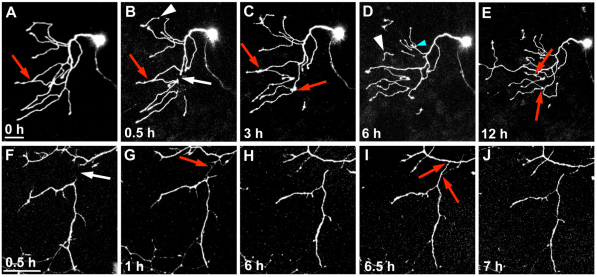
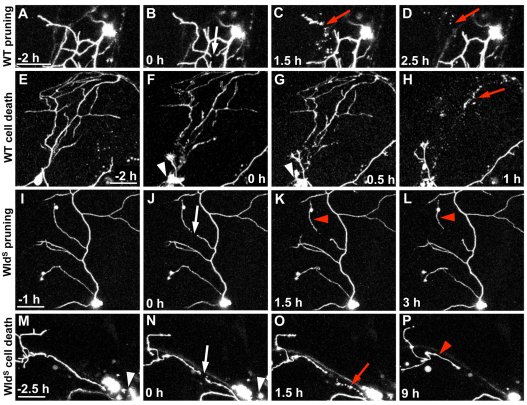
Fragments of injured axons that detach from their cell body break down by the molecularly regulated process of Wallerian degeneration (WD). Although WD resembles local axon degeneration, a common mechanism for refining neuronal structure, several previously examined instances of developmental pruning were unaffected by WD pathways. We used laser axotomy and time-lapse confocal imaging to characterize and compare peripheral sensory axon WD and developmental pruning in live zebrafish larvae. Detached fragments of single injured axon arbors underwent three stereotyped phases of WD: a lag phase, a fragmentation phase and clearance. The lag phase was developmentally regulated, becoming shorter as embryos aged, while the length of the clearance phase increased with the amount of axon debris. Both cell-specific inhibition of ubiquitylation and overexpression of the Wallerian degeneration slow protein (Wld(S)) lengthened the lag phase dramatically, but neither affected fragmentation. Persistent Wld(S)-expressing axon fragments directly repelled regenerating axon branches of their parent arbor, similar to self-repulsion among sister branches of intact arbors. Expression of Wld(S) also disrupted naturally occurring local axon pruning and axon degeneration in spontaneously dying trigeminal neurons: although pieces of Wld(S)-expressing axons were pruned, and some Wld(S)-expressing cells still died during development, in both cases detached axon fragments failed to degenerate. We propose that spontaneously pruned fragments of peripheral sensory axons must be removed by a WD-like mechanism to permit efficient innervation of the epidermis.
Figures
References
-
- Adalbert R., Gillingwater T. H., Haley J. E., Bridge K., Beirowski B., Berek L., Wagner D., Grumme D., Thomson D., Celik A., et al. (2005). A rat model of slow Wallerian degeneration (WldS) with improved preservation of neuromuscular synapses. Eur. J. Neurosci. 21, 271-277 - PubMed
-
- Araki T., Sasaki Y., Milbrandt J. (2004). Increased nuclear NAD biosynthesis and SIRT1 activation prevent axonal degeneration. Science 305, 1010-1013 - PubMed
-
- Awasaki T., Ito K. (2004). Engulfing action of glial cells is required for programmed axon pruning during Drosophila metamorphosis. Curr. Biol. 14, 668-677 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
