

Review
doi: 10.1101/gad.1973010.
Alternative pre-mRNA splicing regulation in cancer: pathways and programs unhinged
Affiliations
- PMID: 21041405
- PMCID: PMC2964746
- DOI: 10.1101/gad.1973010
Item in Clipboard
Review
Alternative pre-mRNA splicing regulation in cancer: pathways and programs unhinged
Genes Dev.
.
Abstract
Alternative splicing of mRNA precursors is a nearly ubiquitous and extremely flexible point of gene control in humans. It provides cells with the opportunity to create protein isoforms of differing, even opposing, functions from a single gene. Cancer cells often take advantage of this flexibility to produce proteins that promote growth and survival. Many of the isoforms produced in this manner are developmentally regulated and are preferentially re-expressed in tumors. Emerging insights into this process indicate that pathways that are frequently deregulated in cancer often play important roles in promoting aberrant splicing, which in turn contributes to all aspects of tumor biology.
Figures
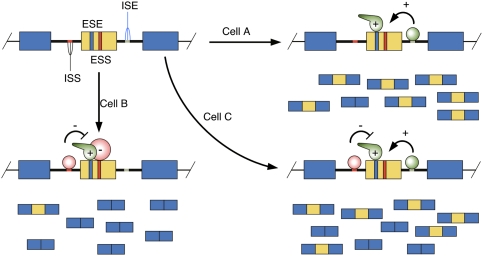
Combinatorial control of splicing by RBPs. In this schematic, a regulated exon (yellow) is flanked by two constitutively spliced exons (blue). Regulatory elements lying within the exon are known as ESEs and ESSs, while intronic regulatory elements are referred to as intronic splicing enhancers (ISEs) and ISSs. The trans-acting factors that bind to these elements can be regulated by changes in intracellular levels, as well as post-translational modifications that affect their cellular localization or activity. The balance of positive- and negative-acting factors present in a given cell determine the extent of regulated exon inclusion.
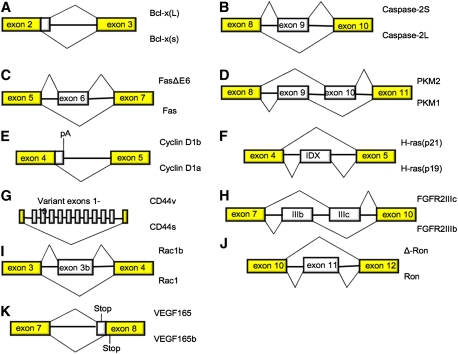
Schematic representation of the AS events discussed in this review. In each case, isoforms that are up-regulated in cancer or that are otherwise shown to have positive effects on growth, survival, or invasive behavior are shown at the top of each diagram.
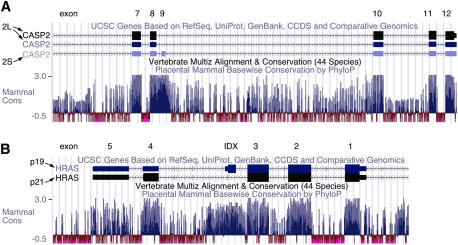
Caspase-2 and H-ras alternative exons lie in highly conserved regions. Images are taken from the University of California at Santa Cruz Genome Browser, using the March 2006 assembly, available at http://genome.ucsc.edu (Kent et al. 2002), with the exon/intron structure of each gene indicated at that top and the mammalian conservation of the corresponding regions indicated below. Conserved regions are indicated by blue bars. (A) For caspase-2, the alternatively spliced exon 9 is highly conserved, and is also flanked by conserved regions, a hallmark of regulated alternatively spliced exons. (B) The IDX exon of H-ras lies within a similarly highly conserved region.
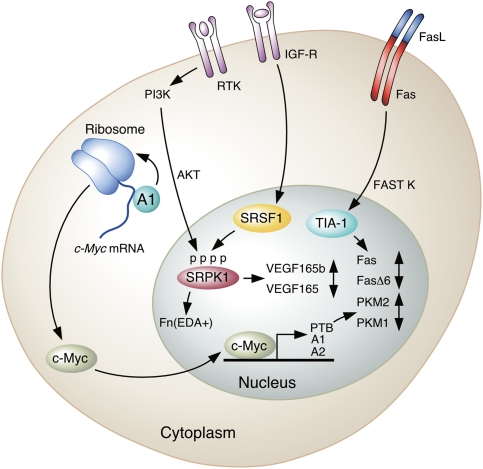
Selected signal transduction pathways that affect AS of transcripts of genes important in cancer. (From top left,) hnRNP A1 promotes translation of c-Myc mRNA. Translated c-Myc protein can promote the transcription of PTB, hnRNP A1, and hnRNP A2, which in turn promote the production of PKM2. The PI3K pathway is activated downstream from ligand binding by receptor tyrosine kinases (RTKs), activating AKT, which phosphorylates SR proteins such as SRSF1, leading to altered splicing of Fn transcripts. IGF1 binding to the IGF receptor (IGF-R) results in activation of the SR protein kinases SRPK1/2, which phosphorylate SRSF1 and alter VEGF165 splicing in favor of the proangiogenesis isoform. Finally, Fas receptor activation by binding to FasL on another cell results in FAST K activation, which in turn results in TIA-1 phosphorylation and inclusion of exon 6 in the Fas transcript.
References
-
- Arch R, Wirth K, Hofmann M, Ponta H, Matzku S, Herrlich P, Zoller M 1992. Participation in normal immune responses of a metastasis-inducing splice variant of CD44. Science 257: 682–685 - PubMed
-
- Babic I, Cherry E, Fujita DJ 2006. SUMO modification of Sam68 enhances its ability to repress cyclin D1 expression and inhibits its ability to induce apoptosis. Oncogene 25: 4955–4964 - PubMed
-
- Barash Y, Calarco JA, Gao W, Pan Q, Wang X, Shai O, Blencowe BJ, Frey BJ 2010. Deciphering the splicing code. Nature 465: 53–59 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources