

Mechanosensitivity of Nav1.5, a voltage-sensitive sodium channel
- PMID: 21041530
- PMCID: PMC3036191
- DOI: 10.1113/jphysiol.2010.199034
Mechanosensitivity of Nav1.5, a voltage-sensitive sodium channel
Abstract
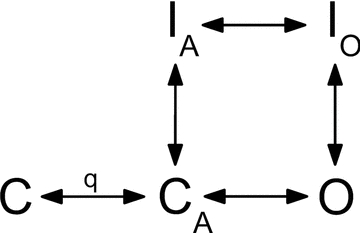
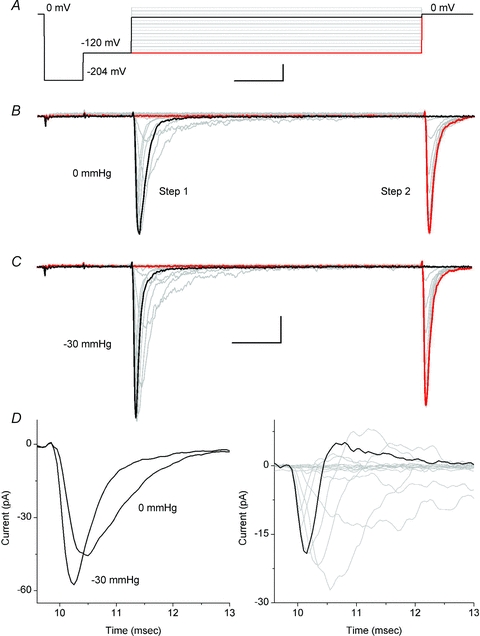
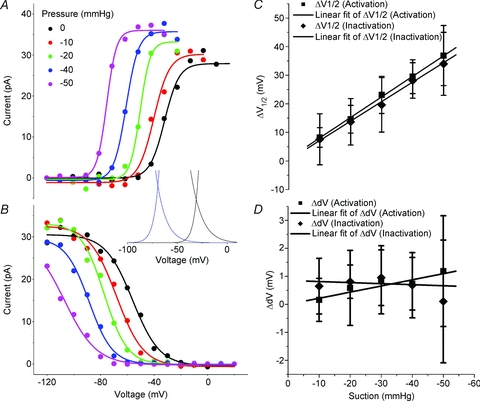
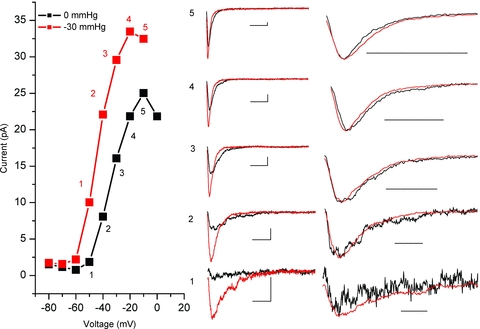
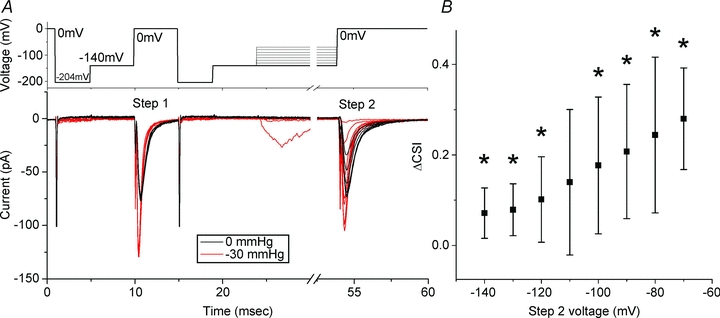
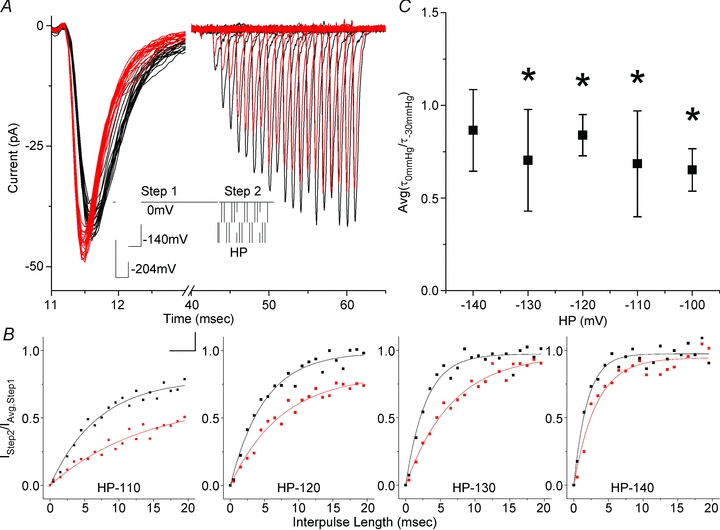
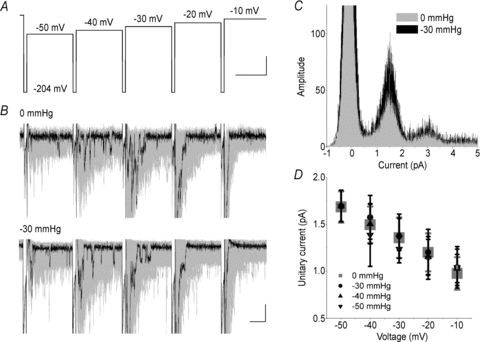
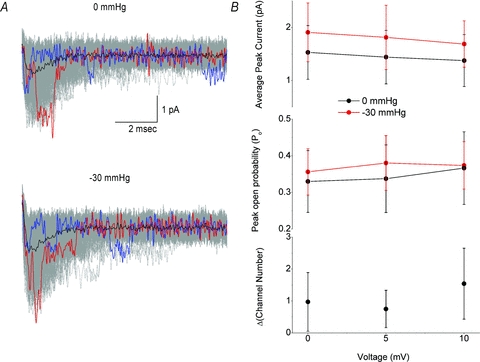
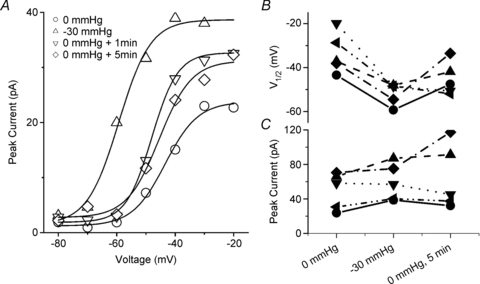
The voltage-sensitive sodium channel Na(v)1.5 (encoded by SCN5A) is expressed in electromechanical organs and is mechanosensitive. This study aimed to determine the mechanosensitive transitions of Na(v)1.5 at the molecular level. Na(v)1.5 was expressed in HEK 293 cells and mechanosensitivity was studied in cell-attached patches. Patch pressure up to -50 mmHg produced increases in current and large hyperpolarizing shifts of voltage dependence with graded shifts of half-activation and half-inactivation voltages (V(1/2)) by ∼0.7 mV mmHg(-1). Voltage dependence shifts affected channel kinetics by a single constant. This suggested that stretch accelerated only one of the activation transitions. Stretch accelerated voltage sensor movement, but not rate constants for gate opening and fast inactivation. Stretch also appeared to stabilize the inactivated states, since recovery from inactivation was slowed with stretch. Unitary conductance and maximum open probability were unaffected by stretch, but peak current was increased due to an increased number of active channels. Stretch effects were partially reversible, but recovery following a single stretch cycle required minutes. These data suggest that mechanical activation of Na(v)1.5 results in dose-dependent voltage dependence shifts of activation and inactivation due to mechanical modulation of the voltage sensors.
Figures
Comment in
-
Sodium channel mechanosensitivity: pay a-tension to voltage sensor movement.J Physiol. 2011 Mar 1;589(Pt 5):1003-4. doi: 10.1113/jphysiol.2011.205880. J Physiol. 2011. PMID: 21486820 Free PMC article. No abstract available.
References
-
- Ackerman MJ. The long QT syndrome: ion channel diseases of the heart. Mayo Clin Proc. 1998;73:250–269. - PubMed
-
- Ackerman MJ, Splawski I, Makielski JC, Tester DJ, Will ML, Timothy KW, Keating MT, Jones G, Chadha M, Burrow CR, Stephens JC, Xu C, Judson R, Curran ME. Spectrum and prevalence of cardiac sodium channel variants among black, white, Asian, and Hispanic individuals: implications for arrhythmogenic susceptibility and Brugada/long QT syndrome genetic testing. Heart Rhythm. 2004;1:600–607. - PubMed
-
- Aldrich RW, Corey DP, Stevens CF. A reinterpretation of mammalian sodium channel gating based on single channel recording. Nature. 1983;306:436–441. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous