

Molecular architecture of the vesicular stomatitis virus RNA polymerase
- PMID: 21041632
- PMCID: PMC2993402
- DOI: 10.1073/pnas.1013559107
Molecular architecture of the vesicular stomatitis virus RNA polymerase
Abstract
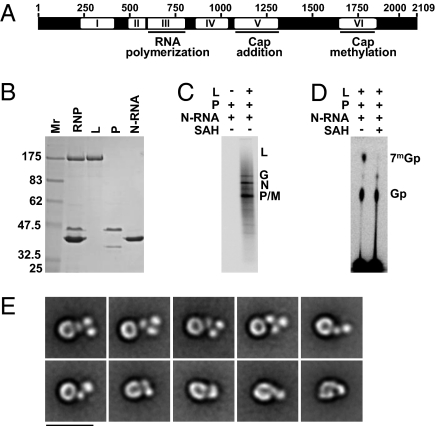
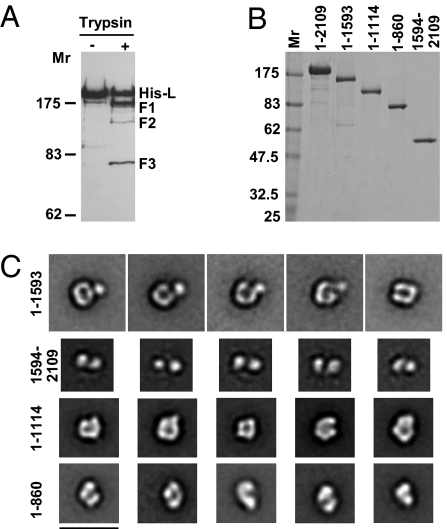
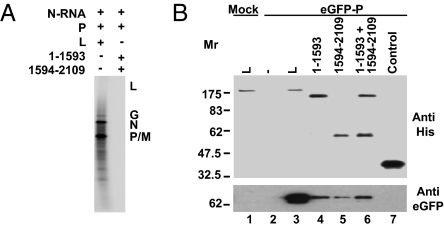
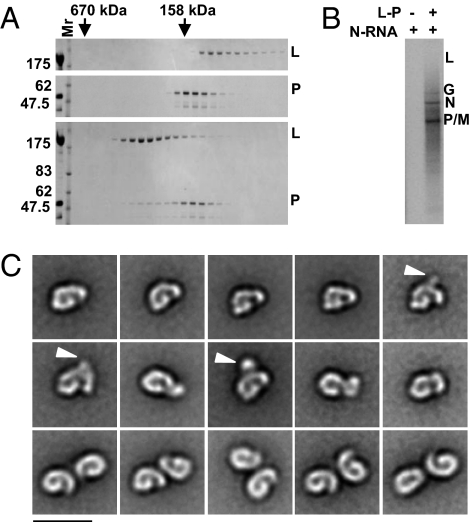
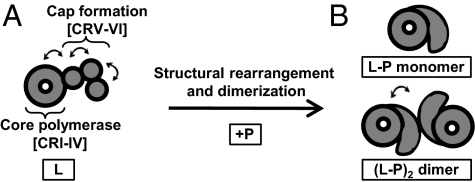
Nonsegmented negative-strand (NNS) RNA viruses initiate infection by delivering into the host cell a highly specialized RNA synthesis machine comprising the genomic RNA completely encapsidated by the viral nucleocapsid protein and associated with the viral polymerase. The catalytic core of this protein-RNA complex is a 250-kDa multifunctional large (L) polymerase protein that contains enzymatic activities for nucleotide polymerization as well as for each step of mRNA cap formation. Working with vesicular stomatitis virus (VSV), a prototype of NNS RNA viruses, we used negative stain electron microscopy (EM) to obtain a molecular view of L, alone and in complex with the viral phosphoprotein (P) cofactor. EM analysis, combined with proteolytic digestion and deletion mapping, revealed the organization of L into a ring domain containing the RNA polymerase and an appendage of three globular domains containing the cap-forming activities. The capping enzyme maps to a globular domain, which is juxtaposed to the ring, and the cap methyltransferase maps to a more distal and flexibly connected globule. Upon P binding, L undergoes a significant rearrangement that may reflect an optimal positioning of its functional domains for transcription. The structural map of L provides new insights into the interrelationship of its various domains, and their rearrangement on P binding that is likely important for RNA synthesis. Because the arrangement of conserved regions involved in catalysis is homologous, the structural insights obtained for VSV L likely extend to all NNS RNA viruses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
