

Nicotinic acid adenine dinucleotide phosphate regulates skeletal muscle differentiation via action at two-pore channels
- PMID: 21041635
- PMCID: PMC2993425
- DOI: 10.1073/pnas.1007381107
Nicotinic acid adenine dinucleotide phosphate regulates skeletal muscle differentiation via action at two-pore channels
Erratum in
- Proc Natl Acad Sci U S A. 2011 Mar 22;108(12):5139
Abstract
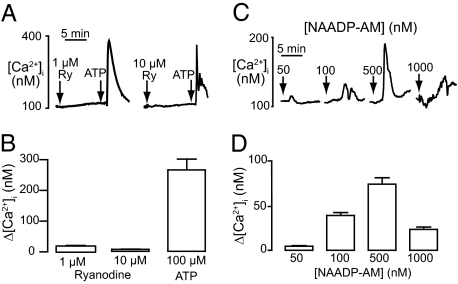
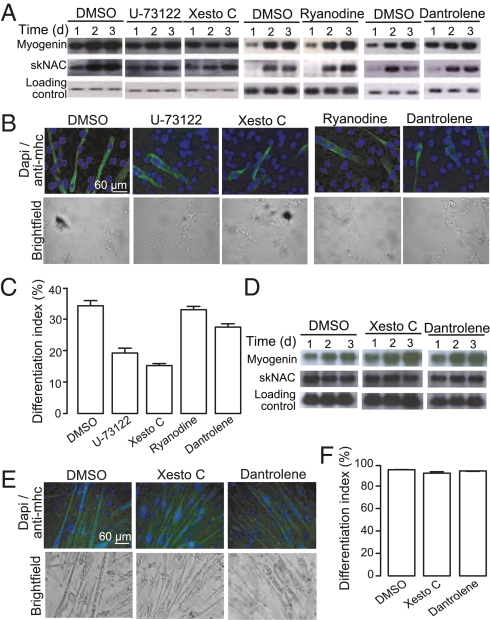
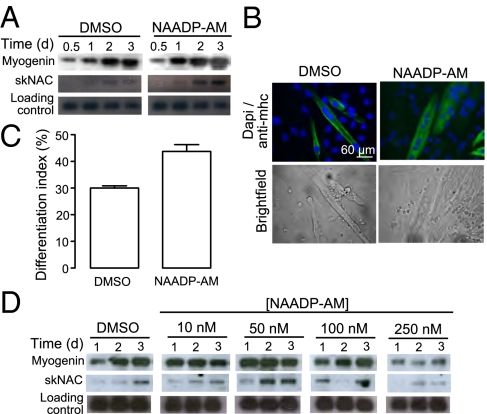
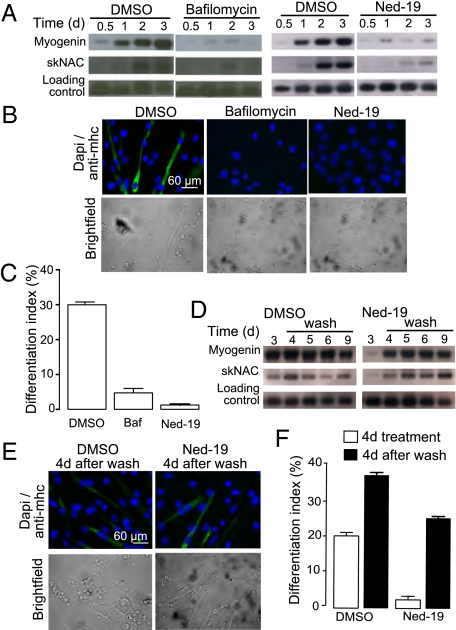
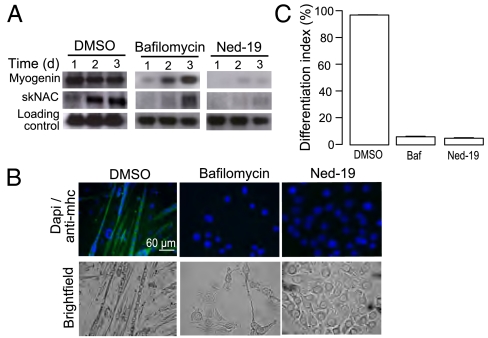
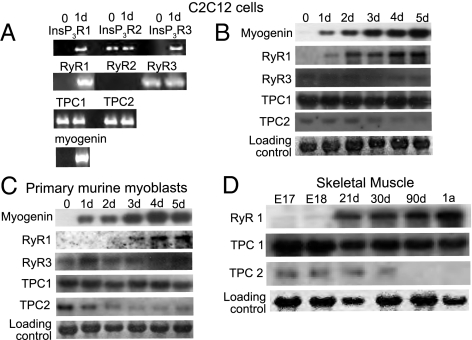
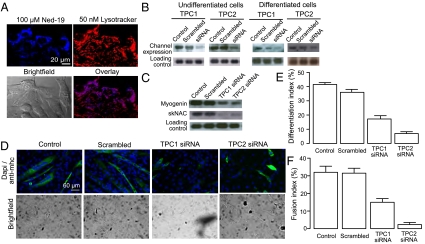
Calcium signaling is essential for the differentiation of many cell types, including skeletal muscle cells, but its mechanisms remain elusive. Here we demonstrate a crucial role for nicotinic acid adenine dinucleotide phosphate (NAADP) signaling in skeletal muscle differentiation. Although the inositol trisphosphate pathway may have a partial role to play in this process, the ryanodine signaling cascade is not involved. In both skeletal muscle precursors and C2C12, cells interfering with NAADP signaling prevented differentiation, whereas promoting NAADP signaling potentiated differentiation. Moreover, siRNA knockdown of two-pore channels, the target of NAADP, attenuated differentiation. The data presented here strongly suggest that in myoblasts, NAADP acts at acidic organelles on the recently discovered two-pore channels to promote differentiation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: Dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003;4:517–529. - PubMed
-
- Ikura M, Osawa M, Ames JB. The role of calcium-binding proteins in the control of transcription: structure to function. Bioessays. 2002;24:625–636. - PubMed
-
- Hogan PG, Chen L, Nardone J, Rao A. Transcriptional regulation by calcium, calcineurin, and NFAT. Genes Dev. 2003;17:2205–2232. - PubMed
-
- Galione A, Ruas M. NAADP receptors. Cell Calcium. 2005;38:273–280. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
