

Mutation of Ser172 in yeast β tubulin induces defects in microtubule dynamics and cell division
- PMID: 21042413
- PMCID: PMC2958848
- DOI: 10.1371/journal.pone.0013553
Mutation of Ser172 in yeast β tubulin induces defects in microtubule dynamics and cell division
Abstract
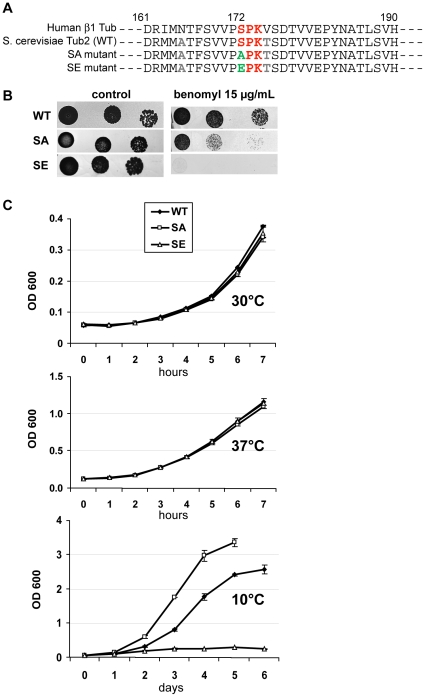
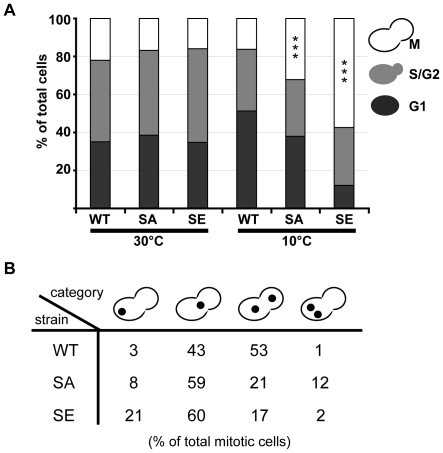
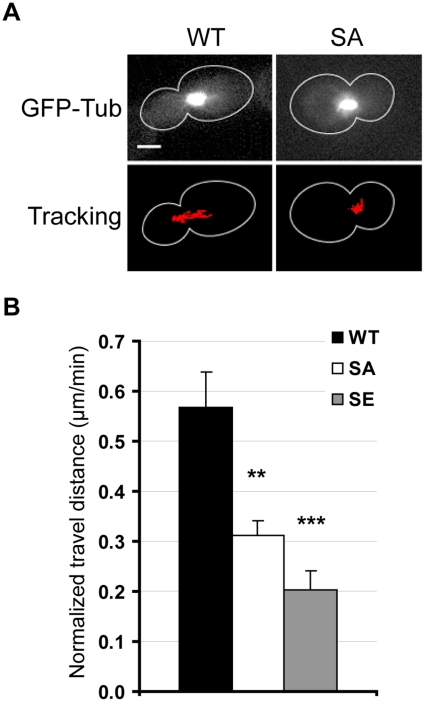
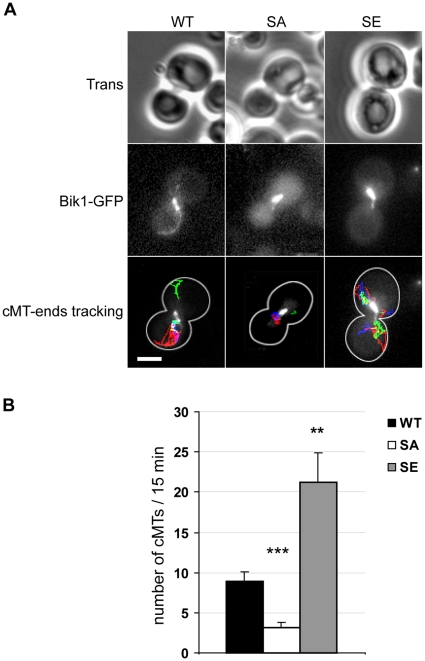
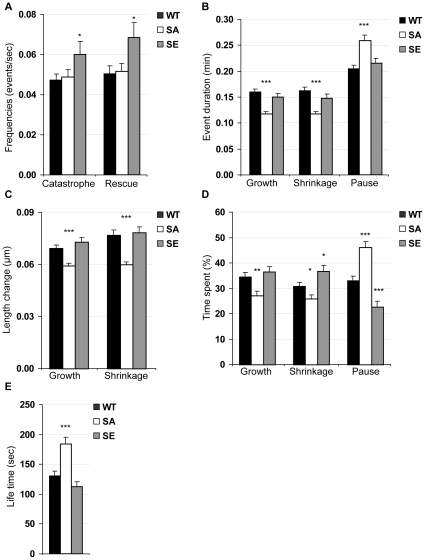
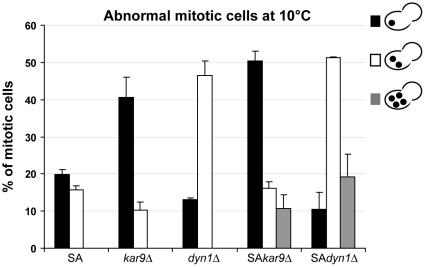
Ser172 of β tubulin is an important residue that is mutated in a human brain disease and phosphorylated by the cyclin-dependent kinase Cdk1 in mammalian cells. To examine the role of this residue, we used the yeast S. cerevisiae as a model and produced two different mutations (S172A and S172E) of the conserved Ser172 in the yeast β tubulin Tub2p. The two mutants showed impaired cell growth on benomyl-containing medium and at cold temperatures, altered microtubule (MT) dynamics, and altered nucleus positioning and segregation. When cytoplasmic MT effectors Dyn1p or Kar9p were deleted in S172A and S172E mutants, cells were viable but presented increased ploidy. Furthermore, the two β tubulin mutations exhibited synthetic lethal interactions with Bik1p, Bim1p or Kar3p, which are effectors of cytoplasmic and spindle MTs. In the absence of Mad2p-dependent spindle checkpoint, both mutations are deleterious. These findings show the importance of Ser172 for the correct function of both cytoplasmic and spindle MTs and for normal cell division.
Conflict of interest statement
Figures
References
-
- Byers B. Cytology of the yeast life cycle. In: Strathern JN, Jones EW, Roach JR, editors. The Molecular Biology of the Yeast Saccharomyces: Life cycle and Inheritance. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory; 1981. pp. 59–96.
-
- Kusch J, Liakopoulos D, Barral Y. Spindle asymmetry: a compass for the cell. Trends Cell Biol. 2003;13:562–569. - PubMed
-
- Huisman SM, Segal M. Cortical capture of microtubules and spindle polarity in budding yeast - where's the catch? J Cell Sci. 2005;118:463–471. - PubMed
-
- Schuyler SC, Pellman D. Search, capture and signal: games microtubules and centrosomes play. J Cell Sci. 2001;114:247–255. - PubMed
-
- Akhmanova A, Hoogenraad CC. Microtubule plus-end-tracking proteins: mechanisms and functions. Curr Opin Cell Biol. 2005;17:47–54. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
