

CLU blocks HDACI-mediated killing of neuroblastoma
- PMID: 21042904
- PMCID: PMC3041908
- DOI: 10.1007/s13277-010-0120-y
CLU blocks HDACI-mediated killing of neuroblastoma
Abstract
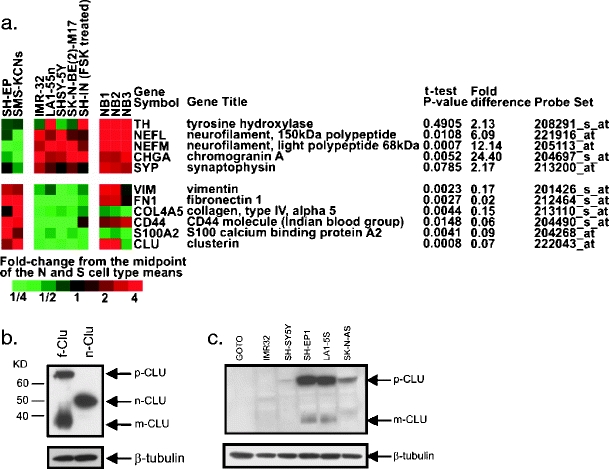
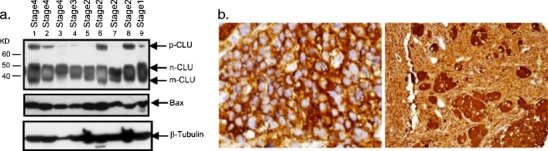
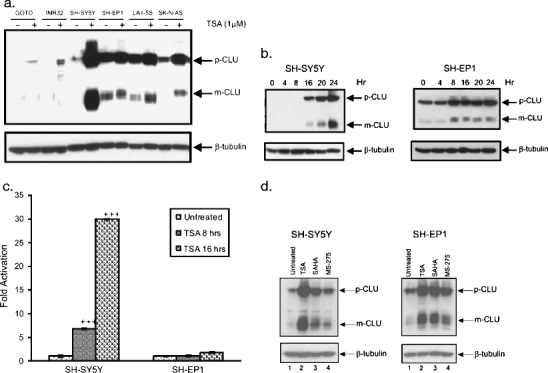
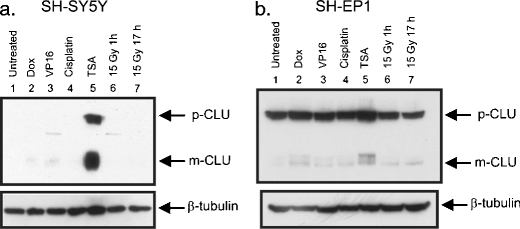
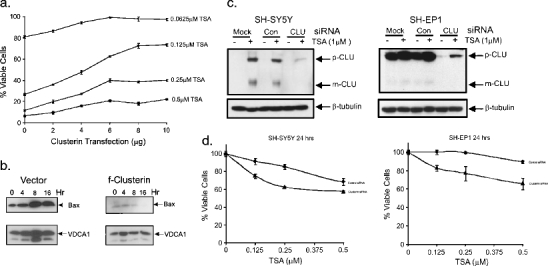
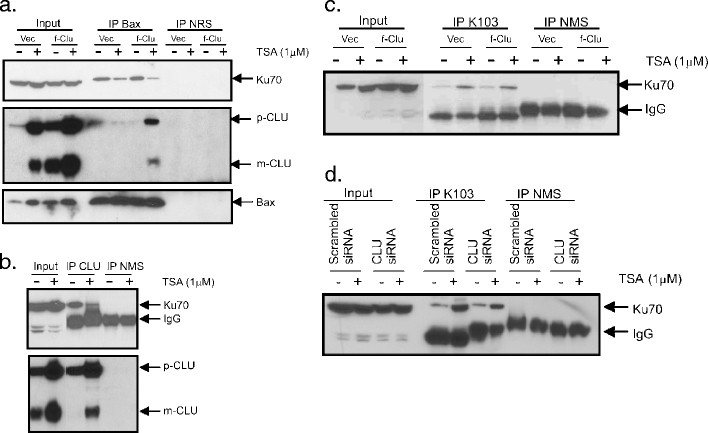
Clusterin is a ubiquitously expressed glycoprotein with multiple binding partners including IL-6, Ku70, and Bax. Clusterin blocks apoptosis by binding to activated Bax and sequestering it in the cytoplasm, thereby preventing Bax from entering mitochondria, releasing cytochrome c, and triggering apoptosis. Because increased clusterin expression correlates with aggressive behavior in tumors, clusterin inhibition might be beneficial in cancer treatment. Our recent findings indicated that, in neuroblastoma cells, cytoplasmic Bax also binds to Ku70; when Ku70 is acetylated, Bax is released and can initiate cell death. Therefore, increasing Ku70 acetylation, such as by using histone deacetylase inhibitors, may be therapeutically useful in promoting cell death in neuroblastoma tumors. Since clusterin, Bax, and Ku70 form a complex, it seemed likely that clusterin would mediate its anti-apoptotic effects by inhibiting Ku70 acetylation and blocking Bax release. Our results, however, demonstrate that while clusterin level does indeed determine the sensitivity of neuroblastoma cells to histone deacetylase inhibitor-induced cell death, it does so without affecting histone deacetylase-inhibitor-induced Ku70 acetylation. Our results suggest that in neuroblastoma, clusterin exerts its anti-apoptotic effects downstream of Ku70 acetylation, likely by directly blocking Bax activation.
Figures
Similar articles
-
HDAC6 deacetylates Ku70 and regulates Ku70-Bax binding in neuroblastoma.Neoplasia. 2011 Aug;13(8):726-34. doi: 10.1593/neo.11558. Neoplasia. 2011. PMID: 21847364 Free PMC article.
-
Ku70 acetylation mediates neuroblastoma cell death induced by histone deacetylase inhibitors.Proc Natl Acad Sci U S A. 2005 Mar 29;102(13):4842-7. doi: 10.1073/pnas.0408351102. Epub 2005 Mar 18. Proc Natl Acad Sci U S A. 2005. PMID: 15778293 Free PMC article.
-
CREB-binding protein is a mediator of neuroblastoma cell death induced by the histone deacetylase inhibitor trichostatin A.Neoplasia. 2007 Jun;9(6):495-503. doi: 10.1593/neo.07262. Neoplasia. 2007. PMID: 17603632 Free PMC article.
-
CLU and colon cancer. The dual face of CLU: from normal to malignant phenotype.Adv Cancer Res. 2009;105:45-61. doi: 10.1016/S0065-230X(09)05003-9. Adv Cancer Res. 2009. PMID: 19879422 Review.
-
Apoptosis Induction byHistone Deacetylase Inhibitors in Cancer Cells: Role of Ku70.Int J Mol Sci. 2019 Mar 30;20(7):1601. doi: 10.3390/ijms20071601. Int J Mol Sci. 2019. PMID: 30935057 Free PMC article. Review.
Cited by
-
Cytoplasmic-translocated Ku70 senses intracellular DNA and mediates interferon-lambda1 induction.Immunology. 2021 Jul;163(3):323-337. doi: 10.1111/imm.13318. Epub 2021 Mar 7. Immunology. 2021. PMID: 33548066 Free PMC article.
-
Se-methylselenocysteine inhibits apoptosis induced by clusterin knockdown in neuroblastoma N2a and SH-SY5Y cell lines.Int J Mol Sci. 2014 Nov 18;15(11):21331-47. doi: 10.3390/ijms151121331. Int J Mol Sci. 2014. PMID: 25411798 Free PMC article.
References
-
- Blaschuk O, Burdzy K, Fritz IB. Purification and characterization of a cell-aggregating factor (clusterin), the major glycoprotein in ram rete testis fluid. J Biol Chem. 1983;258(12):7714–7720. - PubMed
-
- Miyake H, et al. Testosterone-repressed prostate message-2 is an antiapoptotic gene involved in progression to androgen independence in prostate cancer. Cancer Res. 2000;60(1):170–176. - PubMed
-
- Sensibar JA, et al. Prevention of cell death induced by tumor necrosis factor alpha in LNCaP cells by overexpression of sulfated glycoprotein-2 (clusterin) Cancer Res. 1995;55(11):2431–2437. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
