

The basics of brain development
- PMID: 21042938
- PMCID: PMC2989000
- DOI: 10.1007/s11065-010-9148-4
The basics of brain development
Abstract
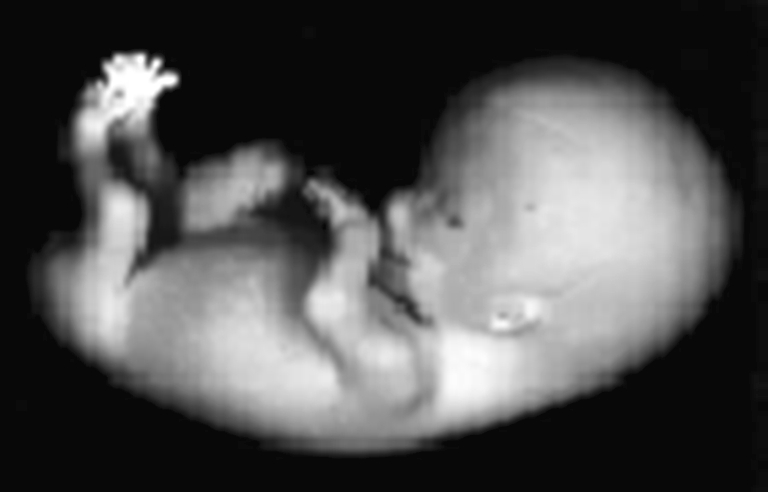
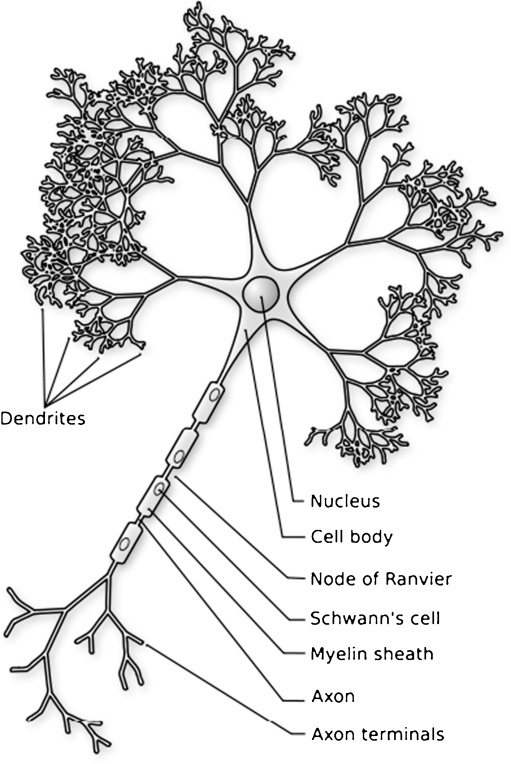
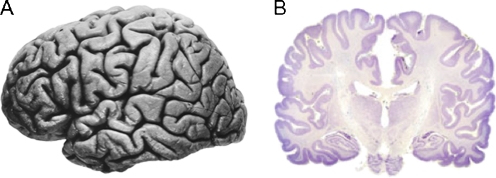
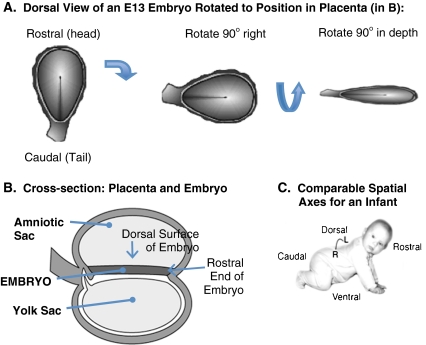
Over the past several decades, significant advances have been made in our understanding of the basic stages and mechanisms of mammalian brain development. Studies elucidating the neurobiology of brain development span the levels of neural organization from the macroanatomic, to the cellular, to the molecular. Together this large body of work provides a picture of brain development as the product of a complex series of dynamic and adaptive processes operating within a highly constrained, genetically organized but constantly changing context. The view of brain development that has emerged from the developmental neurobiology literature presents both challenges and opportunities to psychologists seeking to understand the fundamental processes that underlie social and cognitive development, and the neural systems that mediate them. This chapter is intended to provide an overview of some very basic principles of brain development, drawn from contemporary developmental neurobiology, that may be of use to investigators from a wide range of disciplines.
Figures
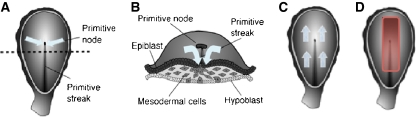
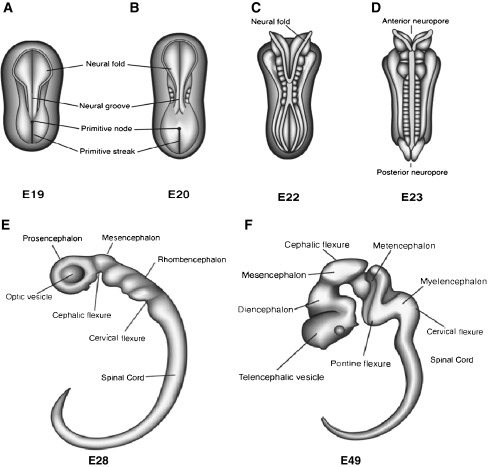
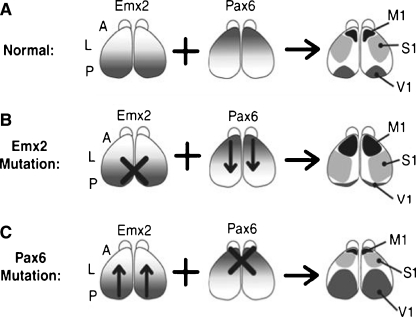
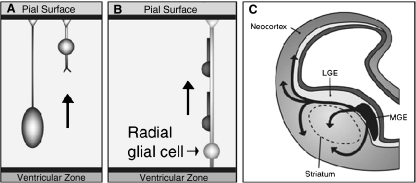
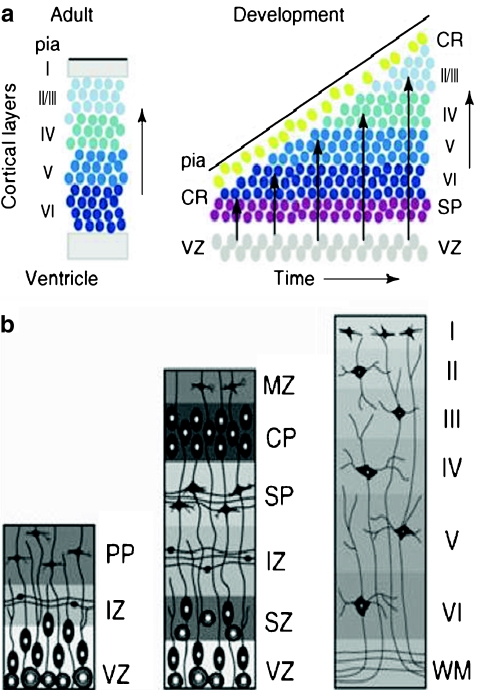
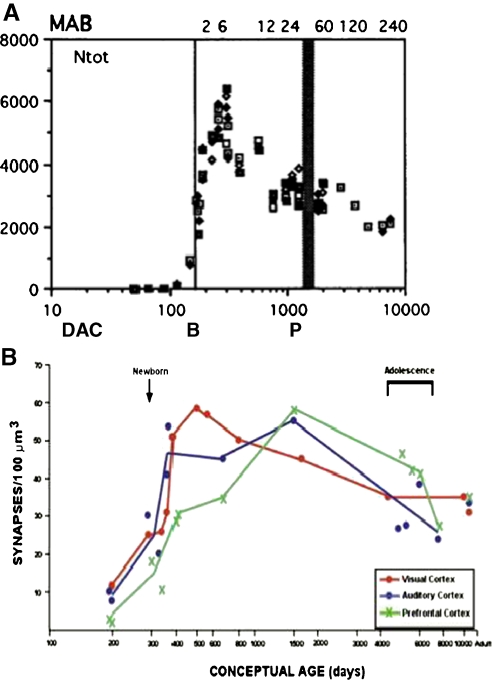
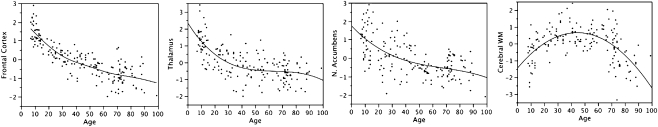
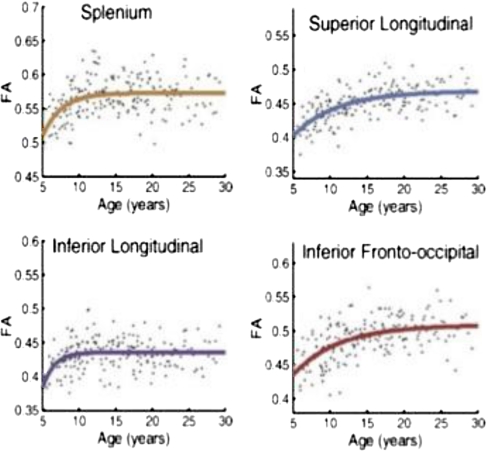
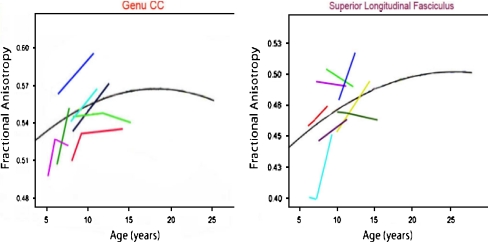
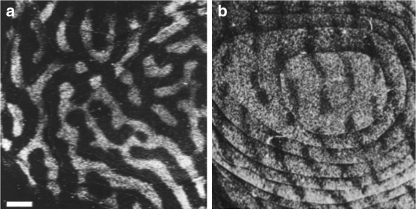
References
-
- Anderson SA, Marin O, et al. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development. 2001;128(3):353–363. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
