

Mechanical forces regulate elastase activity and binding site availability in lung elastin
- PMID: 21044606
- PMCID: PMC2965944
- DOI: 10.1016/j.bpj.2010.09.018
Mechanical forces regulate elastase activity and binding site availability in lung elastin
Abstract
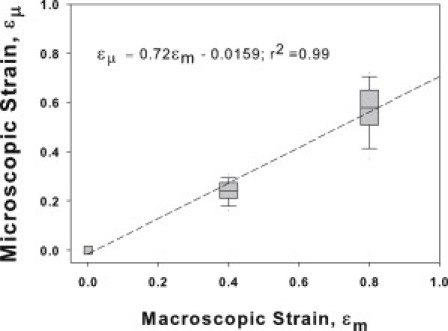
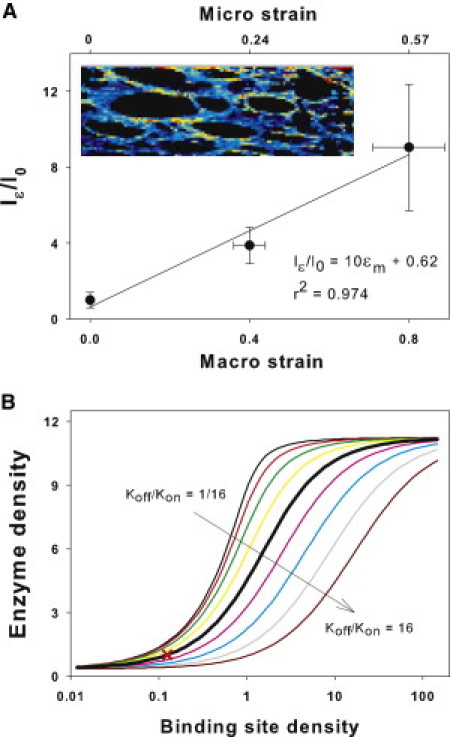
Many fundamental cellular and extracellular processes in the body are mediated by enzymes. At the single molecule level, enzyme activity is influenced by mechanical forces. However, the effects of mechanical forces on the kinetics of enzymatic reactions in complex tissues with intact extracellular matrix (ECM) have not been identified. Here we report that physiologically relevant macroscopic mechanical forces modify enzyme activity at the molecular level in the ECM of the lung parenchyma. Porcine pancreatic elastase (PPE), which binds to and digests elastin, was fluorescently conjugated (f-PPE) and fluorescent recovery after photobleach was used to evaluate the binding kinetics of f-PPE in the alveolar walls of normal mouse lungs. Fluorescent recovery after photobleach indicated that the dissociation rate constant (k(off)) for f-PPE was significantly larger in stretched than in relaxed alveolar walls with a linear relation between k(off) and macroscopic strain. Using a network model of the parenchyma, a linear relation was also found between k(off) and microscopic strain on elastin fibers. Further, the binding pattern of f-PPE suggested that binding sites on elastin unfold with strain. The increased overall reaction rate also resulted in stronger structural breakdown at the level of alveolar walls, as well as accelerated decay of stiffness and decreased failure stress of the ECM at the macroscopic scale. These results suggest an important role for the coupling between mechanical forces and enzyme activity in ECM breakdown and remodeling in development, and during diseases such as pulmonary emphysema or vascular aneurysm. Our findings may also have broader implications because in vivo, enzyme activity in nearly all cellular and extracellular processes takes place in the presence of mechanical forces.
Copyright © 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
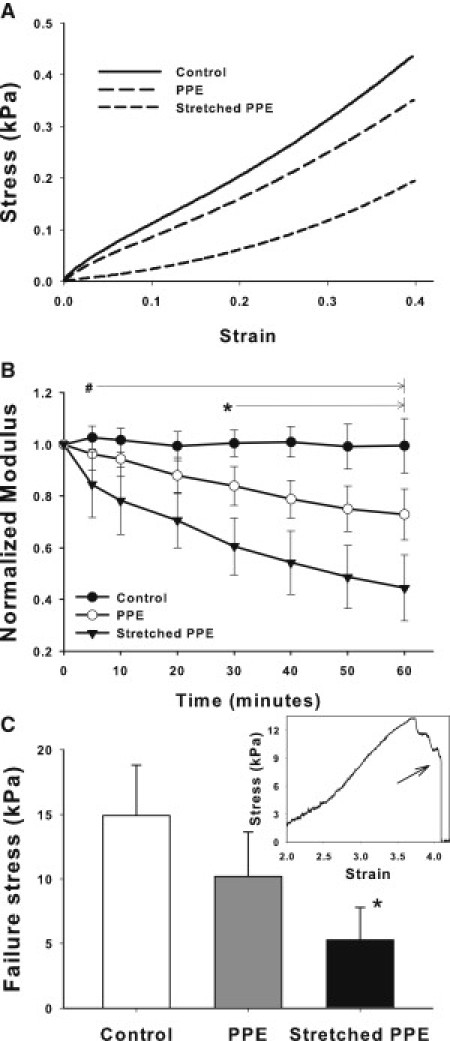
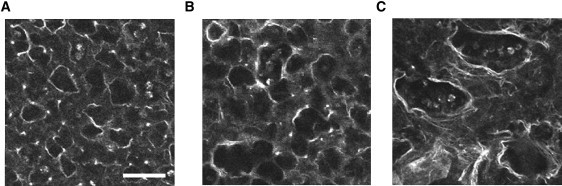
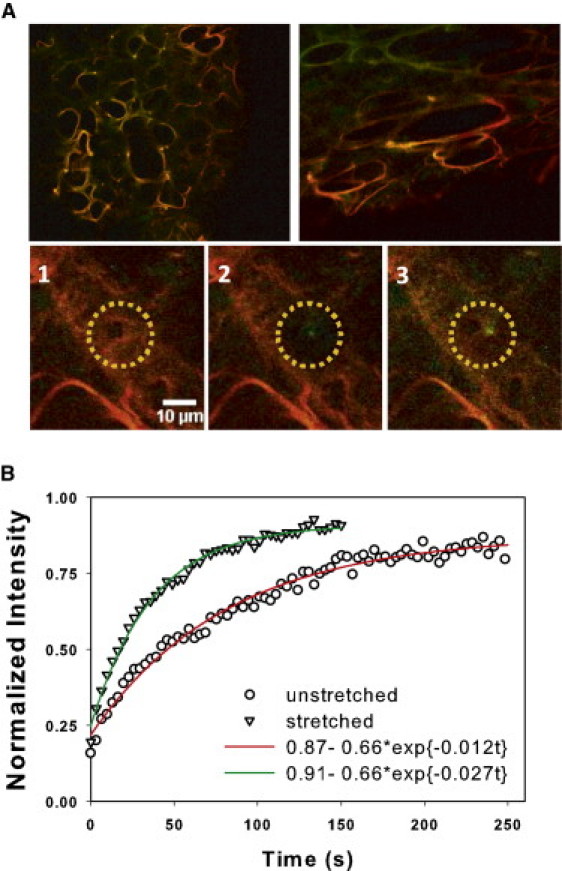
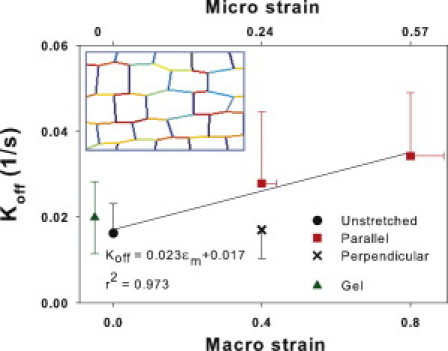
References
-
- Alberts B. Garland Science; New York: 2008. Molecular Biology of the Cell.
-
- Mecham R.P., Broekelmann T.J., Senior R.M. Elastin degradation by matrix metalloproteinases. Cleavage site specificity and mechanisms of elastolysis. J. Biol. Chem. 1997;272:18071–18076. - PubMed
-
- Khan S., Sheetz M.P. Force effects on biochemical kinetics. Annu. Rev. Biochem. 1997;66:785–805. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
