

Nonlinearity and noise at the rod-rod bipolar cell synapse
- PMID: 21047445
- PMCID: PMC3058690
- DOI: 10.1017/S0952523810000301
Nonlinearity and noise at the rod-rod bipolar cell synapse
Abstract
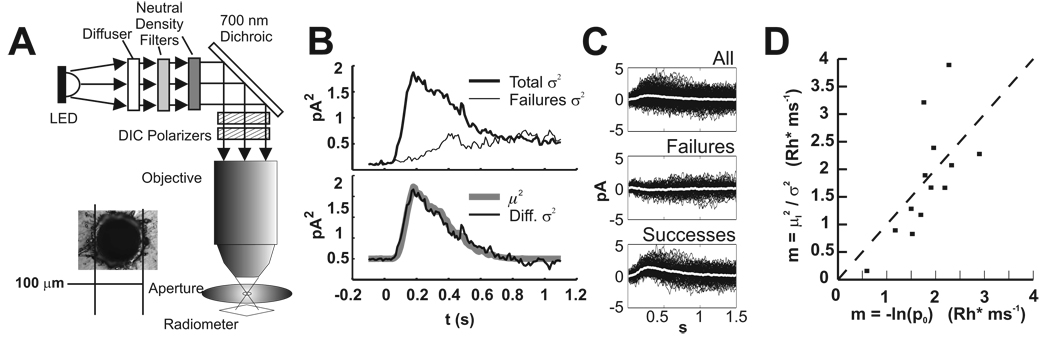
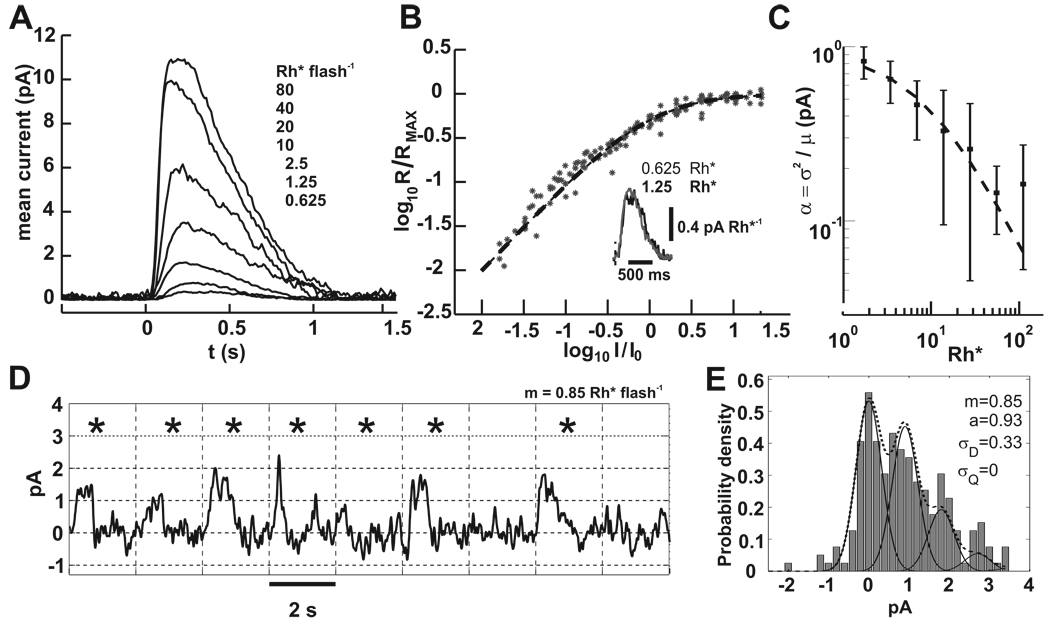
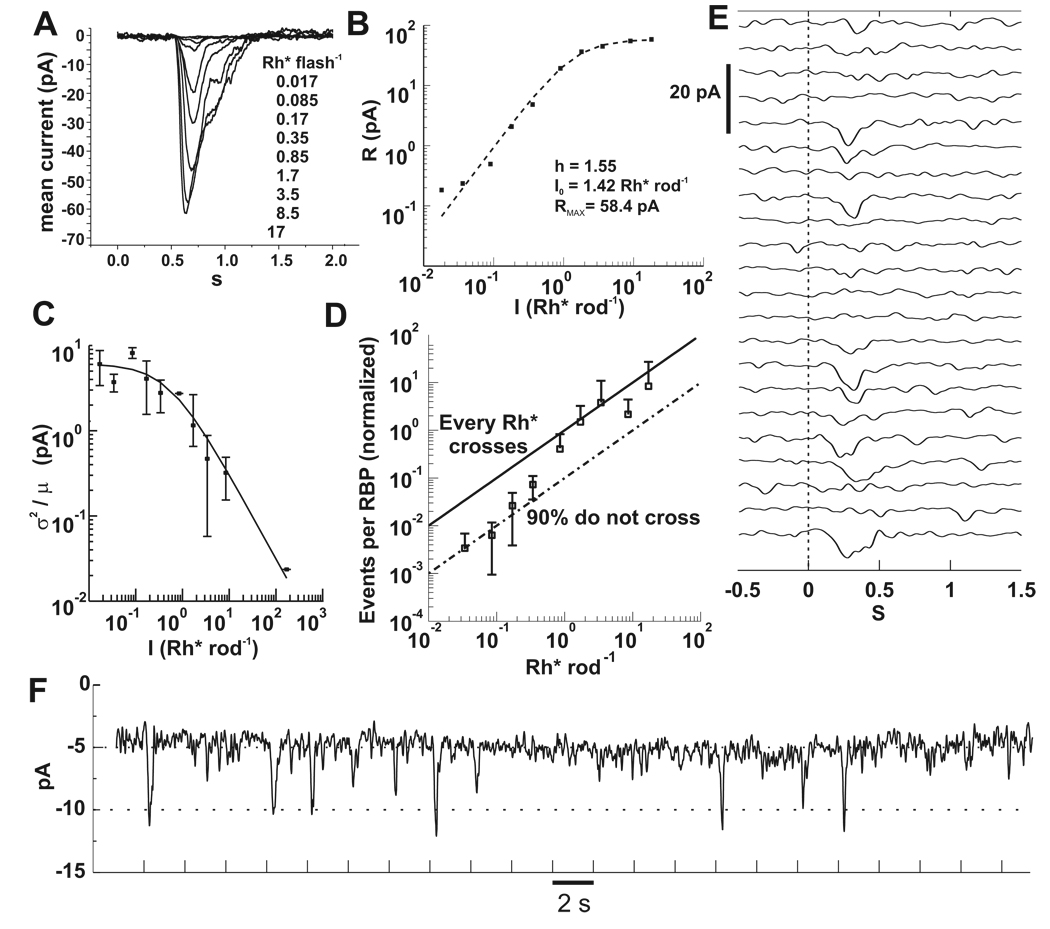
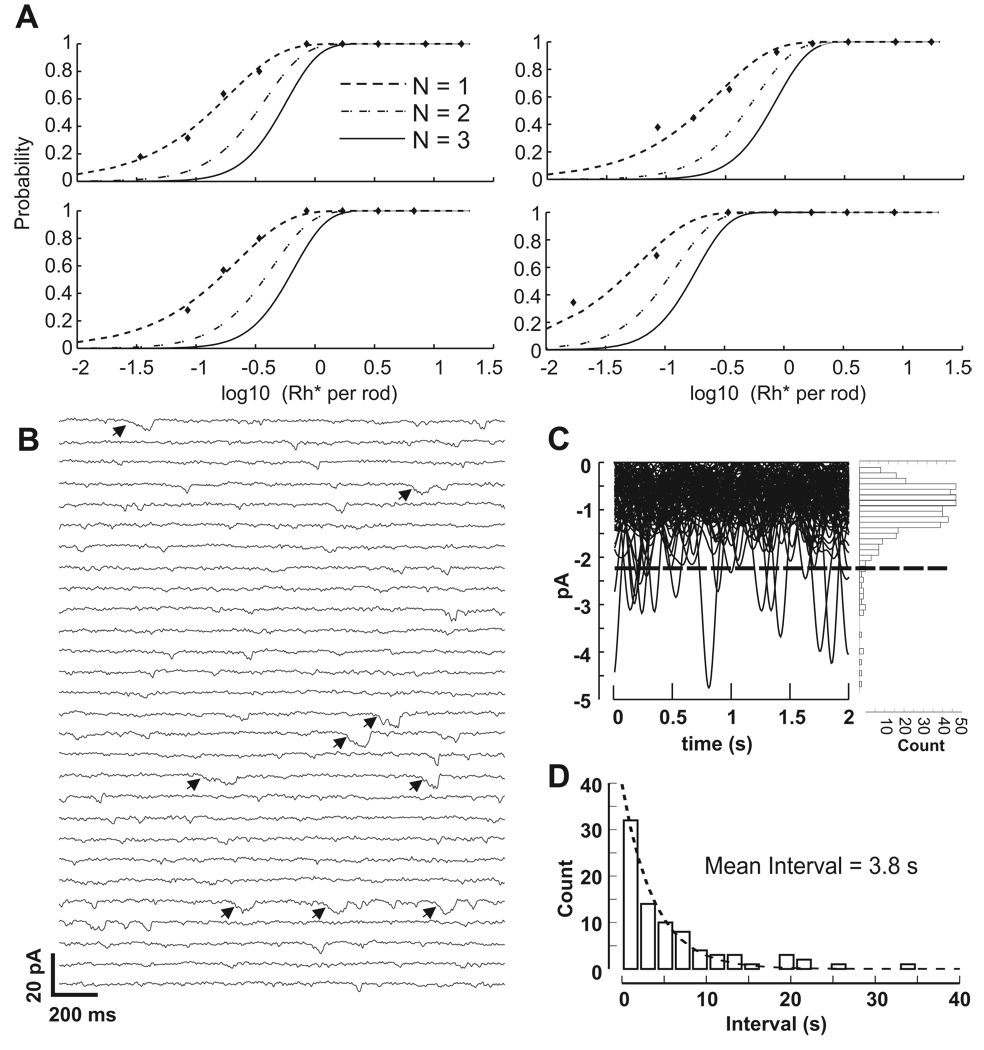
In the retina, rod bipolar (RBP) cells synapse with many rods, and suppression of rod outer segment and synaptic noise is necessary for their detection of rod single-photon responses (SPRs). Depending on the rods' signal-to-noise ratio (SNR), the suppression mechanism will likely eliminate some SPRs as well, resulting in decreased quantum efficiency. We examined this synapse in rabbit, where 100 rods converge onto each RBP. Suction electrode recordings showed that rabbit rod SPRs were difficult to distinguish from noise (independent SNR estimates were 2.3 and 2.8). Nonlinear transmission from rods to RBPs improved response detection (SNR = 8.7), but a large portion of the rod SPRs was discarded. For the dimmest flashes, the loss approached 90%. Despite the high rejection ratio, noise of two distinct types was apparent in the RBP traces: low-amplitude rumblings and discrete events that resembled the SPR. The SPR-like event frequency suggests that they result from thermal isomerizations of rhodopsin, which occurred at the rate 0.033/s/rod. The presence of low-amplitude noise is explained by a sigmoidal input-output relationship at the rod-RBP synapse and the input of noisy rods. The rabbit rod SNR and RBP quantum efficiency are the lowest yet reported, suggesting that the quantum efficiency of the rod-RBP synapse may depend on the SNR in rods. These results point to the possibility that fewer photoisomerizations are discarded for species such as primate, which has a higher rod SNR.
Figures
Similar articles
-
Noise removal at the rod synapse of mammalian retina.Vis Neurosci. 1998 Sep-Oct;15(5):809-21. doi: 10.1017/s0952523898155037. Vis Neurosci. 1998. PMID: 9764523
-
In vivo studies of signaling in rod pathways of the mouse using the electroretinogram.Vision Res. 2004 Dec;44(28):3253-68. doi: 10.1016/j.visres.2004.09.002. Vision Res. 2004. PMID: 15535993
-
Ex Vivo Functional Evaluation of Synaptic Transmission from Rods to Rod Bipolar Cells in Mice.Methods Mol Biol. 2018;1753:203-216. doi: 10.1007/978-1-4939-7720-8_14. Methods Mol Biol. 2018. PMID: 29564791 Free PMC article.
-
Transmission of scotopic signals from the rod to rod-bipolar cell in the mammalian retina.Vision Res. 2004 Dec;44(28):3269-76. doi: 10.1016/j.visres.2004.07.043. Vision Res. 2004. PMID: 15535994 Review.
-
Optimization of single-photon response transmission at the rod-to-rod bipolar synapse.Physiology (Bethesda). 2007 Aug;22:279-86. doi: 10.1152/physiol.00007.2007. Physiology (Bethesda). 2007. PMID: 17699881 Review.
Cited by
-
Regional variation in the organization and connectivity of the first synapse in the primate night vision pathway.iScience. 2023 Oct 2;26(11):108113. doi: 10.1016/j.isci.2023.108113. eCollection 2023 Nov 17. iScience. 2023. PMID: 37915604 Free PMC article.
-
Properties of multivesicular release from mouse rod photoreceptors support transmission of single-photon responses.Elife. 2021 Mar 26;10:e67446. doi: 10.7554/eLife.67446. Elife. 2021. PMID: 33769285 Free PMC article.
-
Elevated cAMP improves signal-to-noise ratio in amphibian rod photoreceptors.J Gen Physiol. 2017 Jul 3;149(7):689-701. doi: 10.1085/jgp.201611744. Epub 2017 Jun 13. J Gen Physiol. 2017. PMID: 28611079 Free PMC article.
-
Voltage-Gated Calcium Channels: Key Players in Sensory Coding in the Retina and the Inner Ear.Physiol Rev. 2018 Oct 1;98(4):2063-2096. doi: 10.1152/physrev.00030.2017. Physiol Rev. 2018. PMID: 30067155 Free PMC article. Review.
-
Visual threshold is set by linear and nonlinear mechanisms in the retina that mitigate noise: how neural circuits in the retina improve the signal-to-noise ratio of the single-photon response.Bioessays. 2011 Jun;33(6):438-47. doi: 10.1002/bies.201100014. Epub 2011 Apr 7. Bioessays. 2011. PMID: 21472740 Free PMC article. Review.
References
-
- Berntson A, Smith RG, Taylor WR. Transmission of single photon signals through a binary synapse in the mammalian retina. Vis Neurosci. 2004;21(5):693–702. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
