

doi: 10.1074/jbc.M110.146738.
Epub 2010 Nov 3.
Mesencephalic astrocyte-derived neurotrophic factor (MANF) has a unique mechanism to rescue apoptotic neurons
Affiliations
- PMID: 21047780
- PMCID: PMC3024763
- DOI: 10.1074/jbc.M110.146738
Item in Clipboard
Mesencephalic astrocyte-derived neurotrophic factor (MANF) has a unique mechanism to rescue apoptotic neurons
J Biol Chem.
.
Abstract
Mesencephalic astrocyte-derived neurotrophic factor (MANF) protects neurons and repairs the Parkinson disease-like symptoms in a rat 6-hydroxydopamine model. We show a three-dimensional solution structure of human MANF that differs drastically from other neurotrophic factors. Remarkably, the C-terminal domain of MANF (C-MANF) is homologous to the SAP domain of Ku70, a well known inhibitor of proapoptotic Bax (Bcl-2-associated X protein). Cellular studies confirm that MANF and C-MANF protect neurons intracellularly as efficiently as Ku70.
Figures

Schematic ribbon presentation of overall fold of MANF and plot of heteronuclear {1H}-15N NOE values. A, structure of FL-MANF is color-coded from N to C terminus starting from blue via green to red. N- and C-terminal domains are connected with flexible linker region (see also supplemental Fig. S1 ). B, the structure of isolated C-MANF is shown. Color-coding of C-MANF is similar to FL-MANF. The structure of C-MANF is composed of three helices, the first helix (α6) is loosely formed and the two consecutive helices (α7 and α8) are found in parallel orientation forming a helix-loop-helix arrangement. Disulfide bridges are shown in magenta. The three-dimensional structure of isolated C-MANF is highly similar to the C-terminal domain of intact full-length MANF. C, shown is a plot of heteronuclear steady-state {1H}-15N NOE values as a function of primary structure. The y axis represents the {1H}-15N NOE values obtained from the ratios of peak intensities in the saturated spectrum and in the unsaturated spectrum. Ratios of <0.6 were ranked as flexible regions. These were found between two domains and at both termini.

Structural comparison of C-MANF with its homologue proteins with Z-score values >4 and r.m.s.d. value <3. A, ribbon presentation of C-MANF in green (PDB code 2KVE). B, C-Ku70 in cyan (PDB code 1JEQ) (25). C, SAP domain of FYVE RING finger protein Sakura in purple (PDB code 1Y02) (26). D, sequences aligned according to structural comparison. Secondary structure elements (helices) of the proteins are labeled with sequences as yellow boxes. Sequences are numbered according to maturated full-length proteins. VPMLKE motif of Ku70 and corresponding region of C-MANF and SAP domain of FYVE RING finger protein Sakura are marked with blue boxes, and the regions aligned to the CXXC motif of C-MANF are shown as green boxes. According to the structural alignment, the first helix in the C-Ku70 and FYVE RING finger protein Sakura is better defined than the corresponding helix α6 in C-MANF. Disulfide bond of C-MANF formed by CXXC motif marked in yellow on the structure. The first cysteine of SAP domain of FYVE RING finger protein Sakura is replaced by threonine. The loop connecting helices α7 and α8 in C-MANF is two residues shorter in C-Ku70, but the disulfide bond can be aligned against the backbone of Ku70. The VPMLKE motif (residues 578–583) of Ku70 is marked in blue ribbons, and the corresponding regions of C-MANF and FYVE RING finger protein Sakura are marked in red and pink ribbons, respectively. The side chains of conserved lysines are marked as red, and conserved hydrophobic amino acids are localized into the α-helical regions (gray side chains and boxes) as a part of the hydrophobic core.
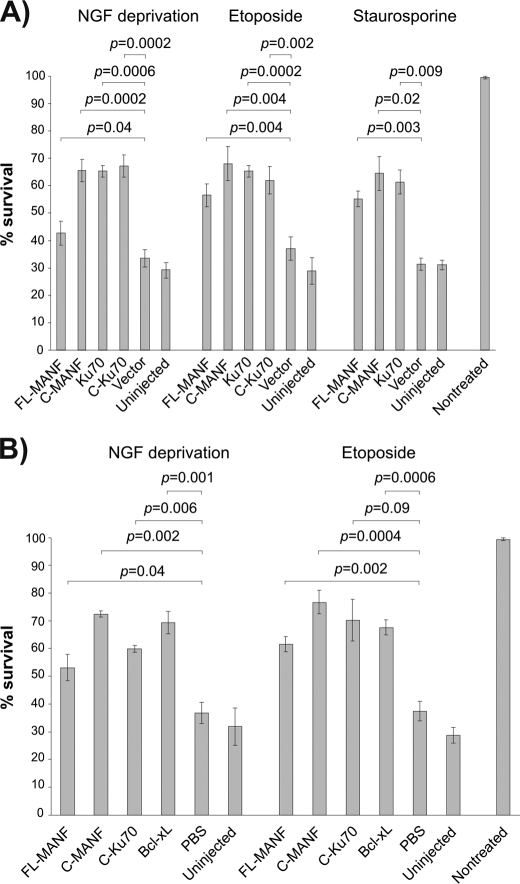
Overexpressed FL-MANF and C-MANF protect apoptotic neurons. Newborn mouse SCG neurons were cultured for 5–6 days with NGF, and then microinjected with the indicated expression plasmids (A) or recombinant proteins (B) and either deprived of NGF or treated with etoposide (30 μm ) or staurosporine (200 nm ) for 3 days. Live injected neurons were then counted and expressed as percent of initially injected neurons. The mean ± S.E. of eight (A, NGF deprivation, etoposide), three (staurosporine), or four (B) independent experiments is shown. Data of each experimental group were compared with control plasmid pcDNA3 (vector) in A, or control PBS in B, by one-way analysis of variance and post hoc Dunnett's t test. The null hypothesis was rejected at p < 0.05.
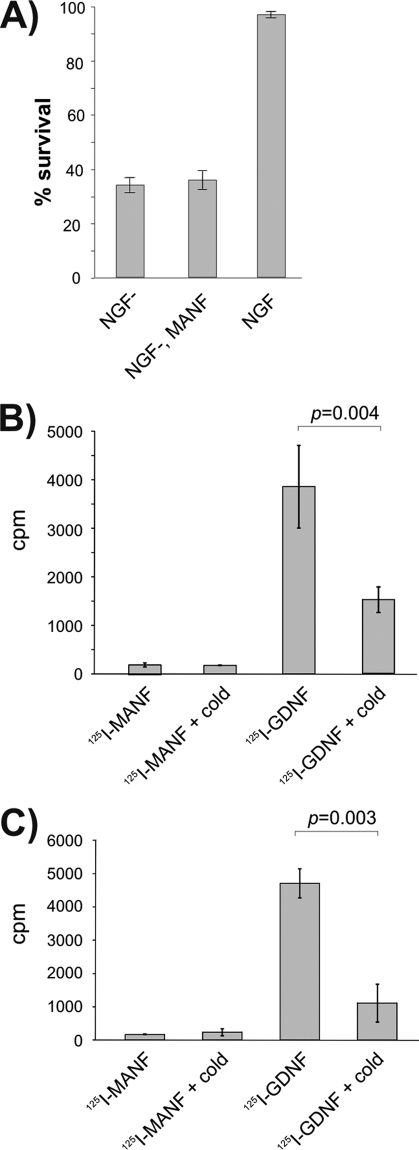
Exogeneous MANF does not affect SCG neurons in vitro. A, MANF protein in the culture medium did not protect SCG neurons. Newborn mouse SCG neurons were cultured with NGF for 5 days, and then NGF was removed (NGF−) and replaced or not replaced with recombinant MANF at 100 ng/ml. Living neurons were counted 3 days later and expressed as percent of initially counted neurons. The mean ± S.E. of four independent experiments is shown. B and C, 125I-labeled MANF is not bound or taken up by the cultured SCG neurons. 125I- MANF or 125I-GDNF was bound to the cultured SCG neurons in the presence or absence of 200-fold excess of respective unlabeled factors (cold). Bound (B) or internalized (C) radioactivity was measured. Shown are the cpm values from the representative experiments (means of three parallels ± S.E.). The means were compared by Student's t test, and the null hypothesis was rejected at p < 0.05. Qualitatively same results were obtained from three independent experiments for both assays.
References
-
- Dauer W., Przedborski S. (2003) Neuron 39, 889–909 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
