

Chromatin boundaries, insulators, and long-range interactions in the nucleus
- PMID: 21047907
- PMCID: PMC9373008
- DOI: 10.1101/sqb.2010.75.006
Chromatin boundaries, insulators, and long-range interactions in the nucleus
Abstract
Within the genome, expressed genes marked by "open" chromatin are often adjacent to silent, heterochromatic regions. There are also regions containing neighboring active genes with different programs of expression. In both cases, DNA sequence elements may function as insulators, either providing barriers that prevent the incursion of heterochromatic signals into open domains or acting to block inappropriate contact between the enhancer of one gene and the promoter of another. The mechanisms associated with insulation are diverse: Enhancer-blocking insulation is largely associated with the ability to stabilize the formation of loop domains within the nucleus. Barrier insulation is often associated with the ability to block propagation of silencing histone modifications. Here, we provide examples of both kinds of insulator action, derived initially from studies of the compound insulator element at the 5' end of the chicken β-globin locus. Such elements appear to have more general regulatory roles in the genome that have been exploited to provide insulator function where necessary to demarcate separate domains within the nucleus.
Figures


Similar articles
-
Chromatin domains, insulators, and the regulation of gene expression.Biochim Biophys Acta. 2012 Jul;1819(7):644-51. doi: 10.1016/j.bbagrm.2012.01.016. Epub 2012 Feb 2. Biochim Biophys Acta. 2012. PMID: 22326678 Free PMC article. Review.
-
Chromatin insulator elements: establishing barriers to set heterochromatin boundaries.Epigenomics. 2012 Feb;4(1):67-80. doi: 10.2217/epi.11.112. Epigenomics. 2012. PMID: 22332659 Review.
-
A chromatin insulator-like element in the herpes simplex virus type 1 latency-associated transcript region binds CCCTC-binding factor and displays enhancer-blocking and silencing activities.J Virol. 2006 Mar;80(5):2358-68. doi: 10.1128/JVI.80.5.2358-2368.2006. J Virol. 2006. PMID: 16474142 Free PMC article.
-
Insulators and domains of gene expression.Curr Opin Genet Dev. 2016 Apr;37:17-26. doi: 10.1016/j.gde.2015.11.009. Epub 2016 Jan 20. Curr Opin Genet Dev. 2016. PMID: 26802288 Review.
-
Multiple CTCF sites cooperate with each other to maintain a TAD for enhancer-promoter interaction in the β-globin locus.FASEB J. 2021 Aug;35(8):e21768. doi: 10.1096/fj.202100105RR. FASEB J. 2021. PMID: 34245617
Cited by
-
Development of gene therapy for thalassemia.Cold Spring Harb Perspect Med. 2012 Nov 1;2(11):a011833. doi: 10.1101/cshperspect.a011833. Cold Spring Harb Perspect Med. 2012. PMID: 23125203 Free PMC article. Review.
-
Telomeres, tethers and trypanosomes.Nucleus. 2012 Nov-Dec;3(6):478-86. doi: 10.4161/nucl.22167. Epub 2012 Sep 19. Nucleus. 2012. PMID: 22992703 Free PMC article.
-
A single heterochromatin boundary element imposes position-independent antisilencing activity in Saccharomyces cerevisiae minichromosomes.PLoS One. 2011;6(9):e24835. doi: 10.1371/journal.pone.0024835. Epub 2011 Sep 16. PLoS One. 2011. PMID: 21949764 Free PMC article.
-
Entropy gives rise to topologically associating domains.Nucleic Acids Res. 2016 Jul 8;44(12):5540-9. doi: 10.1093/nar/gkw510. Epub 2016 Jun 2. Nucleic Acids Res. 2016. PMID: 27257057 Free PMC article.
-
Regulatory Elements in Vectors for Efficient Generation of Cell Lines Producing Target Proteins.Acta Naturae. 2015 Jul-Sep;7(3):15-26. Acta Naturae. 2015. PMID: 26483956 Free PMC article.
References
-
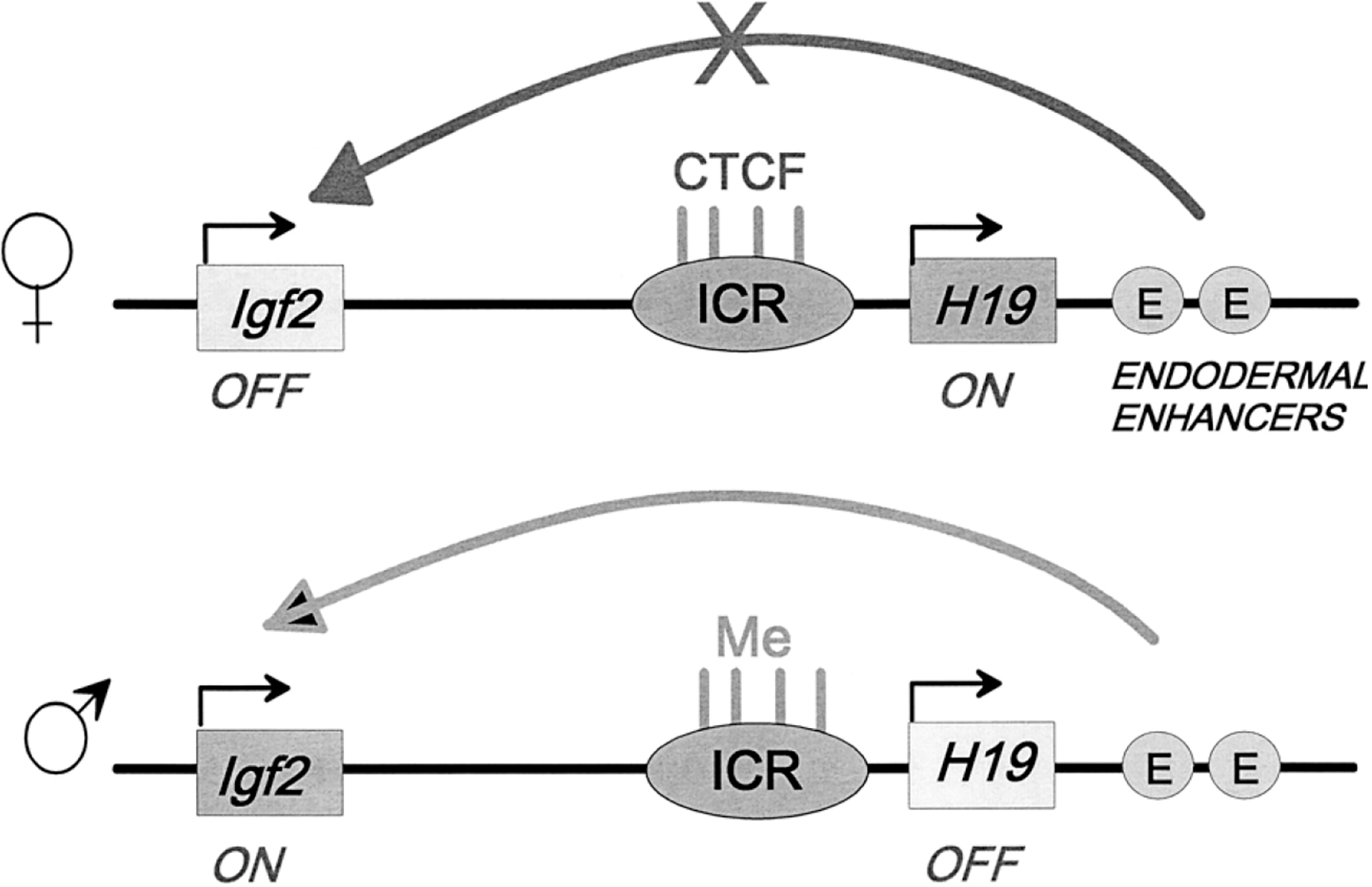
- Bell AC, Felsenfeld G. 2000. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature 405: 482–485. - PubMed
-
- Bell AC, West AG, Felsenfeld G. 1999. The protein CTCF is required for the enhancer blocking activity of vertebrate insulators. Cell 98: 387–396. - PubMed
-
- Cai H, Levine M. 1995. Modulation of enhancer-promoter interactions by insulators in the Drosophila embryo. Nature 376: 533–536. - PubMed
-
- Chung JH, Whiteley M, Felsenfeld G. 1993. A 5′ element of the chicken β-globin domain serves as an insulator in human erythroid cells and protects against position effect in Drosophila. Cell 74: 505–514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources