

GAD67-GFP+ neurons in the Nucleus of Roller: a possible source of inhibitory input to hypoglossal motoneurons. I. Morphology and firing properties
- PMID: 21047932
- PMCID: PMC3023366
- DOI: 10.1152/jn.00493.2010
GAD67-GFP+ neurons in the Nucleus of Roller: a possible source of inhibitory input to hypoglossal motoneurons. I. Morphology and firing properties
Abstract
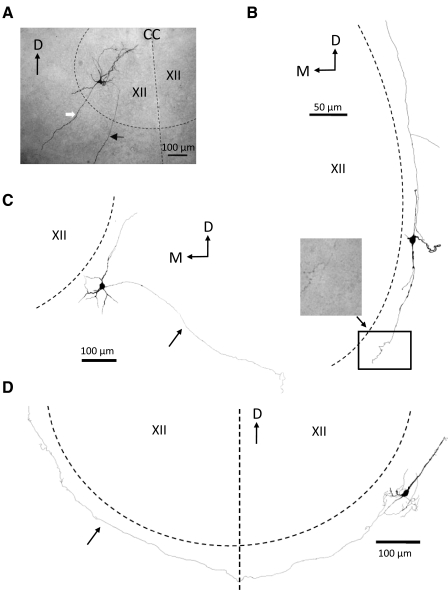
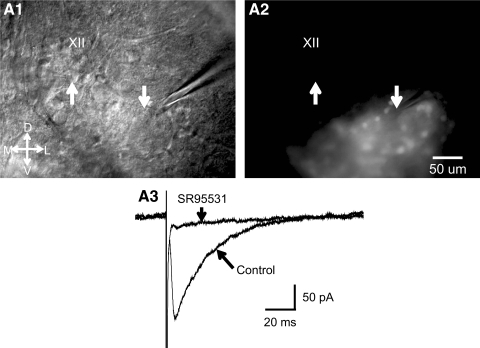
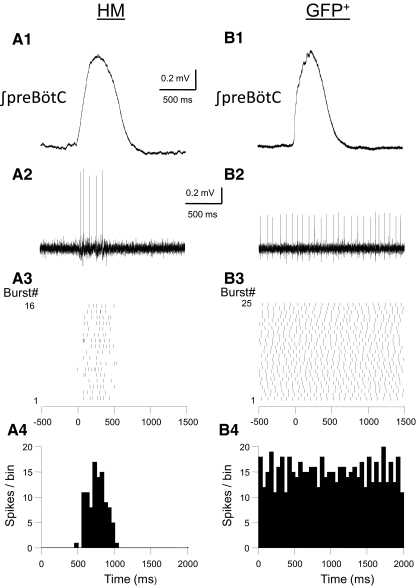
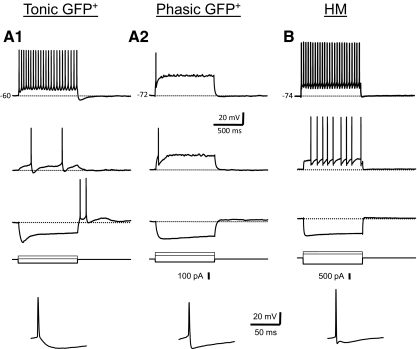
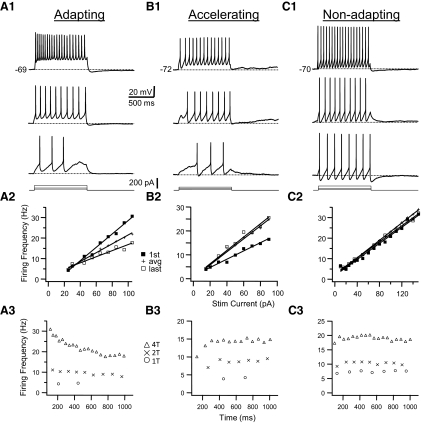
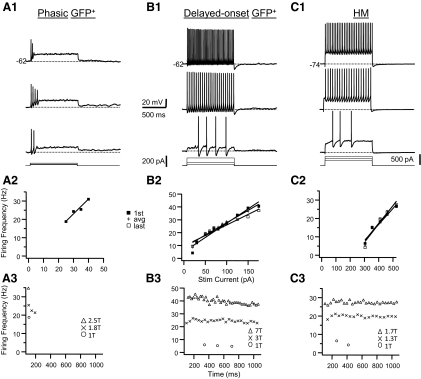
In this study we examined the electrophysiological and morphological properties of inhibitory neurons located just ventrolateral to the hypoglossal motor (XII) nucleus in the Nucleus of Roller (NR). In vitro experiments were performed on medullary slices derived from postnatal day 5 (P5) to P15 GAD67-GFP knock-in mouse pups. on cell recordings from GFP+ cells in NR in rhythmic slices revealed that these neurons are spontaneously active, although their spiking activity does not exhibit inspiratory phase modulation. Morphologically, GFP+ cells were bi- or multipolar cells with small- to medium-sized cell bodies and small dendritic trees that were often oriented parallel to the border of the XII nucleus. GFP+ cells were classified as either tonic or phasic based on their firing responses to depolarizing step current stimulation in whole cell current clamp. Tonic GFP+ cells fired a regular train of action potentials (APs) throughout the duration of the pulse and often showed rebound spikes after a hyperpolarizing step. In contrast, phasic GFP+ neurons did not fire throughout the depolarizing current step but instead fired fewer than four APs at the onset of the pulse or fired multiple APs, but only after a marked delay. Phasic cells had a significantly smaller input resistance and shorter membrane time constant than tonic GFP+ cells. In addition, phasic GFP+ cells differed from tonic cells in the shape and time course of their spike afterpotentials, the minimum firing frequency at threshold current amplitude, and the slope of their current-frequency relationship. These results suggest that GABAergic neurons in the NR are morphologically and electrophysiologically heterogeneous cells that could provide tonic inhibitory synaptic input to HMs.
Figures

Similar articles
-
GAD67-GFP+ neurons in the Nucleus of Roller. II. Subthreshold and firing resonance properties.J Neurophysiol. 2011 Jan;105(1):249-78. doi: 10.1152/jn.00492.2010. Epub 2010 Nov 3. J Neurophysiol. 2011. PMID: 21047931 Free PMC article.
-
Properties of GABAergic neurons in the rostral solitary tract nucleus in mice.J Neurophysiol. 2010 Jun;103(6):3205-18. doi: 10.1152/jn.00971.2009. Epub 2010 Apr 7. J Neurophysiol. 2010. PMID: 20375246 Free PMC article.
-
Electrophysiological and morphological characteristics of GABAergic respiratory neurons in the mouse pre-Bötzinger complex.Eur J Neurosci. 2006 Feb;23(3):667-74. doi: 10.1111/j.1460-9568.2006.04591.x. Eur J Neurosci. 2006. PMID: 16487148
-
GABAergic neurons in inferior colliculus of the GAD67-GFP knock-in mouse: electrophysiological and morphological properties.Neurosci Res. 2005 Apr;51(4):475-92. doi: 10.1016/j.neures.2004.12.019. Neurosci Res. 2005. PMID: 15740810
-
Morphological and electrophysiological properties of GABAergic and non-GABAergic cells in the deep cerebellar nuclei.J Neurophysiol. 2007 Jan;97(1):901-11. doi: 10.1152/jn.00974.2006. Epub 2006 Nov 8. J Neurophysiol. 2007. PMID: 17093116
Cited by
-
P2Y1 receptor-mediated potentiation of inspiratory motor output in neonatal rat in vitro.J Physiol. 2014 Jul 15;592(14):3089-111. doi: 10.1113/jphysiol.2013.268136. Epub 2014 May 30. J Physiol. 2014. PMID: 24879869 Free PMC article.
-
Developmental changes in the morphology of mouse hypoglossal motor neurons.Brain Struct Funct. 2016 Sep;221(7):3755-86. doi: 10.1007/s00429-015-1130-8. Epub 2015 Oct 17. Brain Struct Funct. 2016. PMID: 26476929 Free PMC article.
-
Functional up-regulation of the M-current by retigabine contrasts hyperexcitability and excitotoxicity on rat hypoglossal motoneurons.J Physiol. 2018 Jul;596(13):2611-2629. doi: 10.1113/JP275906. Epub 2018 May 30. J Physiol. 2018. PMID: 29736957 Free PMC article.
-
Respiratory related control of hypoglossal motoneurons--knowing what we do not know.Respir Physiol Neurobiol. 2011 Oct 15;179(1):43-7. doi: 10.1016/j.resp.2011.06.023. Epub 2011 Jul 2. Respir Physiol Neurobiol. 2011. PMID: 21741499 Free PMC article. Review.
-
Lack of an endogenous GABAA receptor-mediated tonic current in hypoglossal motoneurons.J Physiol. 2012 Jul 1;590(13):2965-76. doi: 10.1113/jphysiol.2012.231944. Epub 2012 Apr 10. J Physiol. 2012. PMID: 22495589 Free PMC article.
References
-
- Acuna-Goycolea C, Tamamaki N, Yanagawa Y, Obata K, van den Pol AN. Mechanisms of neuropeptide Y, peptide YY, and pancreatic polypeptide inhibition of identified green fluorescent protein-expressing GABA neurons in the hypothalamic neuroendocrine arcuate nucleus. J Neurosci 25: 7406–7419, 2005 - PMC - PubMed
-
- Aldes LD, Chronister RB, Marco LA. Distribution of glutamic acid decarboxylase and gamma-aminobutyric acid in the hypoglossal nucleus in the rat. J Neurosci Res 19: 343–348, 1988 - PubMed
-
- Altschuler SM, Bao X, Miselis RR. Dendritic architecture of hypoglossal motoneurons projecting to extrinsic tongue musculature in the rat. J Comp Neurol 342: 538–550, 1994 - PubMed
-
- Bou-Flores C, Berger AJ. Gap junctions and inhibitory synapses modulate inspiratory motoneuron synchronization. J Neurophysiol 85: 1543–1551, 2001 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous