

Biomechanical analysis of gait adaptation in the nematode Caenorhabditis elegans
- PMID: 21048086
- PMCID: PMC2996704
- DOI: 10.1073/pnas.1003016107
Biomechanical analysis of gait adaptation in the nematode Caenorhabditis elegans
Abstract
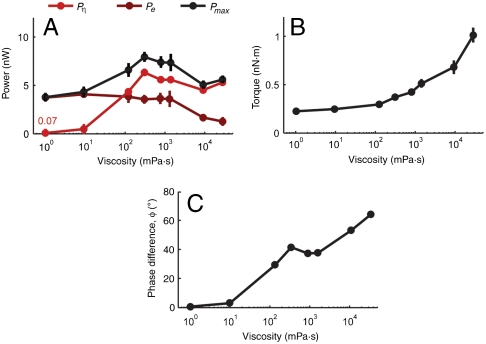
To navigate different environments, an animal must be able to adapt its locomotory gait to its physical surroundings. The nematode Caenorhabditis elegans, between swimming in water and crawling on surfaces, adapts its locomotory gait to surroundings that impose approximately 10,000-fold differences in mechanical resistance. Here we investigate this feat by studying the undulatory movements of C. elegans in Newtonian fluids spanning nearly five orders of magnitude in viscosity. In these fluids, the worm undulatory gait varies continuously with changes in external load: As load increases, both wavelength and frequency of undulation decrease. We also quantify the internal viscoelastic properties of the worm's body and their role in locomotory dynamics. We incorporate muscle activity, internal load, and external load into a biomechanical model of locomotion and show that (i) muscle power is nearly constant across changes in locomotory gait, and (ii) the onset of gait adaptation occurs as external load becomes comparable to internal load. During the swimming gait, which is evoked by small external loads, muscle power is primarily devoted to bending the worm's elastic body. During the crawling gait, evoked by large external loads, comparable muscle power is used to drive the external load and the elastic body. Our results suggest that C. elegans locomotory gait continuously adapts to external mechanical load in order to maintain propulsive thrust.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
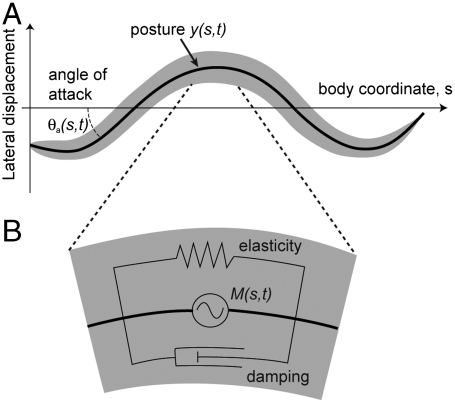
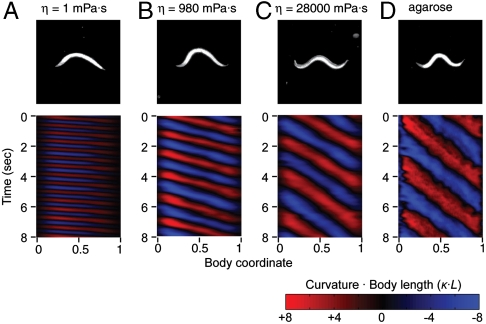
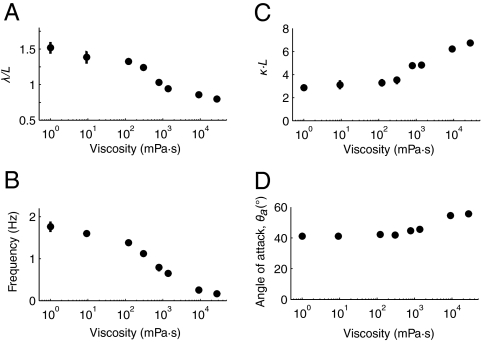
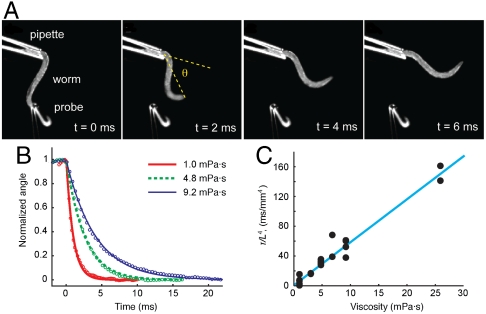
References
-
- White JG, Southgate E, Thomson JN, Brenner S. The structure of the nervous-system of the nematode Caenorhabditis-elegans. Philos T Roy Soc B. 1986;314:1–340. - PubMed
-
- Von Stetina SE, Treinin M, Miller DM., 3rd The motor circuit. Int Rev Neurobiol. 2006;69:125–67. - PubMed
-
- Croll NA. The Behaviour of Nematodes: Their Activity, Senses and Responses. London: Edward Arnold; 1970.
-
- Karbowski J, et al. Conservation rules, their breakdown, and optimality in Caenorhabditis sinusoidal locomotion. J Theor Biol. 2006;242:652–69. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
