

Dynamic plasticity of axons within a cutaneous milieu
- PMID: 21048132
- PMCID: PMC6633624
- DOI: 10.1523/JNEUROSCI.2919-10.2010
Dynamic plasticity of axons within a cutaneous milieu
Abstract
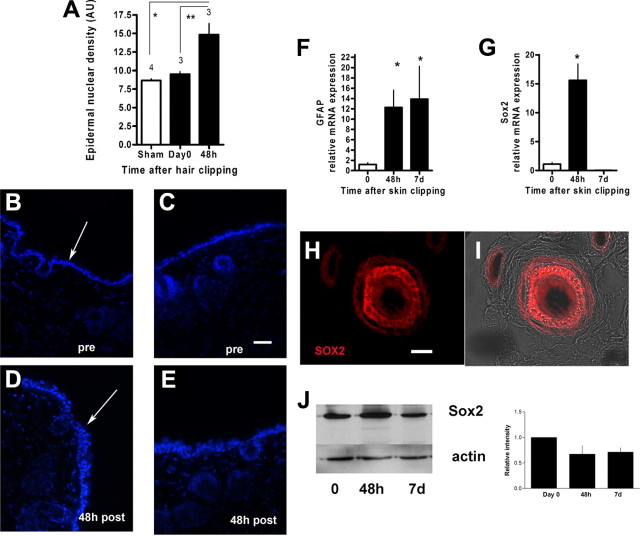
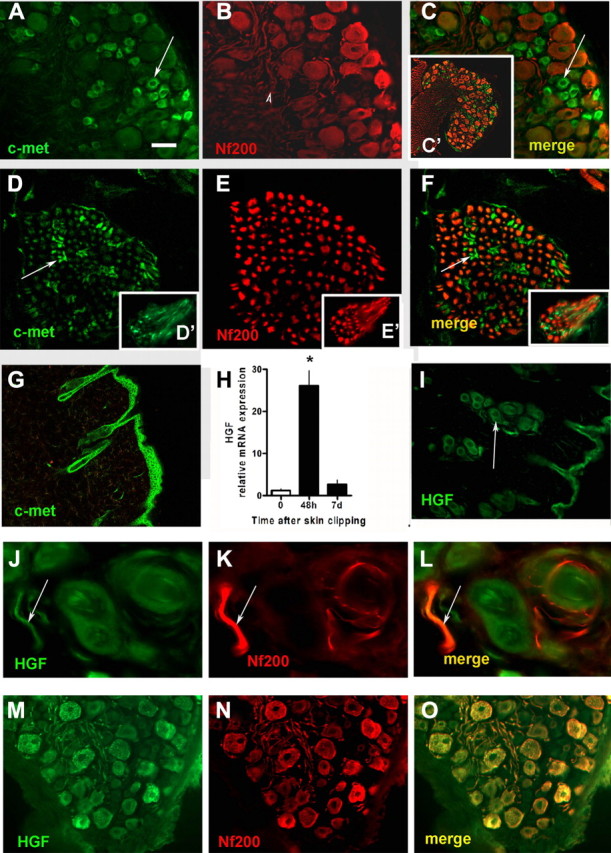
The skin is a repository of sensory axons immersed within the turnover of epidermal, follicular, and dermal cellular constituents. We show that epidermal and perifollicular axons within intact hairy skin of mice possess a remarkable dynamic plasticity linked to their microenvironment. For example, the majority of epidermal axons express the growth protein GAP43. Unexpectedly, we induced new cutaneous axogenesis by simple and noninvasive hair clipping, a response linked to a series of changes in their cutaneous neighbors. In thy-1 YFP transgenic mice with fluorescent axons, superficial epidermal and perifollicular cells newly acquired YFP, indicating diffuse activation by clipping despite the absence of skin injury. At 48 h after clipping, this activation was accompanied by a rise in the number of epidermal cells, transient rises in mRNA of Sox2, a marker of follicular stem cells, and a rise in mRNA of glial fibrillary acidic protein, a marker of glial cells. Axons responded with rises in their numbers in the epidermis and around dermal hair follicles. Linking these responses were early, large, and selective rises in hepatic growth factor (HGF) mRNA, with its protein identified in epidermal cells, perifollicular cells, and sensory axons. Moreover, these elements also expressed the HGF receptor c-Met, especially in small caliber sensory neurons. Finally, we identified concurrent rises in Rac1 activation, a downstream target of ligated c-Met. Together, these results confirm critical linkages between sensory axons and their cutaneous milieu. We believe that the plasticity is provoked by follicular-originating cutaneous activation with HGF and Rac1 signaling, allowing cross talk and axonal remodeling.
Figures




Similar articles
-
Developmental timing of hair follicle and dorsal skin innervation in mice.J Comp Neurol. 2002 Jun 17;448(1):28-52. doi: 10.1002/cne.10212. J Comp Neurol. 2002. PMID: 12012374
-
Selective Sensory Axon Reinnervation and TRPV1 Activation.Mol Neurobiol. 2019 Oct;56(10):7144-7158. doi: 10.1007/s12035-019-1574-6. Epub 2019 Apr 15. Mol Neurobiol. 2019. PMID: 30989631
-
Local insulin and the rapid regrowth of diabetic epidermal axons.Neurobiol Dis. 2011 Aug;43(2):414-21. doi: 10.1016/j.nbd.2011.04.012. Epub 2011 Apr 17. Neurobiol Dis. 2011. PMID: 21530660
-
Diversification and specialization of touch receptors in skin.Cold Spring Harb Perspect Med. 2014 Jun 2;4(6):a013656. doi: 10.1101/cshperspect.a013656. Cold Spring Harb Perspect Med. 2014. PMID: 24890830 Free PMC article. Review.
-
Hair Follicle Transplantation for Wound Repair.Adv Wound Care (New Rochelle). 2021 Mar;10(3):153-163. doi: 10.1089/wound.2019.1139. Epub 2020 Jun 10. Adv Wound Care (New Rochelle). 2021. PMID: 32522101 Free PMC article. Review.
Cited by
-
Repeated monitoring of corneal nerves by confocal microscopy as an index of peripheral neuropathy in type-1 diabetic rodents and the effects of topical insulin.J Peripher Nerv Syst. 2013 Dec;18(4):306-15. doi: 10.1111/jns5.12044. J Peripher Nerv Syst. 2013. PMID: 24147903 Free PMC article.
-
Patterns of cutaneous nerve fibre loss and regeneration in type 2 diabetes with painful and painless polyneuropathy.Diabetologia. 2017 Dec;60(12):2495-2503. doi: 10.1007/s00125-017-4438-5. Epub 2017 Sep 15. Diabetologia. 2017. PMID: 28914336
-
Homeostatic pruning and activity of epidermal nerves are dysregulated in barrier-impaired skin during chronic itch development.Sci Rep. 2019 Jun 13;9(1):8625. doi: 10.1038/s41598-019-44866-0. Sci Rep. 2019. PMID: 31197234 Free PMC article.
-
Gene therapy for diabetic peripheral neuropathy: A randomized, placebo-controlled phase III study of VM202, a plasmid DNA encoding human hepatocyte growth factor.Clin Transl Sci. 2021 May;14(3):1176-1184. doi: 10.1111/cts.12977. Epub 2021 Feb 2. Clin Transl Sci. 2021. PMID: 33465273 Free PMC article. Clinical Trial.
-
Nerve growth factor acts through the TrkA receptor to protect sensory neurons from the damaging effects of the HIV-1 viral protein, Vpr.Neuroscience. 2013 Nov 12;252:512-25. doi: 10.1016/j.neuroscience.2013.07.046. Epub 2013 Jul 30. Neuroscience. 2013. PMID: 23912036 Free PMC article.
References
-
- Biernaskie JA, McKenzie IA, Toma JG, Miller FD. Isolation of skin-derived precursors (SKPs) and differentiation and enrichment of their Schwann cell progeny. Nat Protoc. 2006;1:2803–2812. - PubMed
-
- Botchkarev VA, Paus R. Molecular biology of hair morphogenesis: development and cycling. J Exp Zool B Mol Dev Evol. 2003;298:164–180. - PubMed
-
- Botchkarev VA, Eichmüller S, Johansson O, Paus R. Hair cycle-dependent plasticity of skin and hair follicle innervation in normal murine skin. J Comp Neurol. 1997;386:379–395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous