

Differential short-term plasticity at convergent inhibitory synapses to the substantia nigra pars reticulata
- PMID: 21048144
- PMCID: PMC6633647
- DOI: 10.1523/JNEUROSCI.3895-10.2010
Differential short-term plasticity at convergent inhibitory synapses to the substantia nigra pars reticulata
Abstract
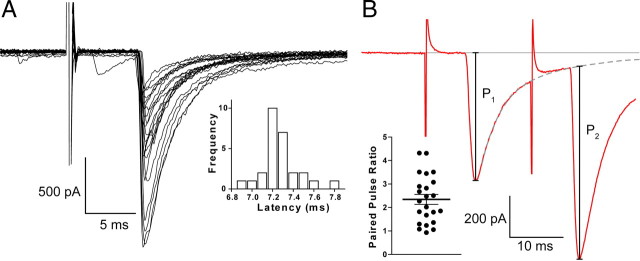
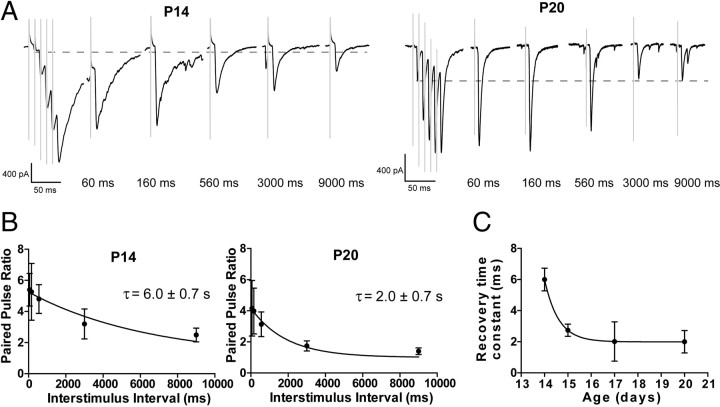
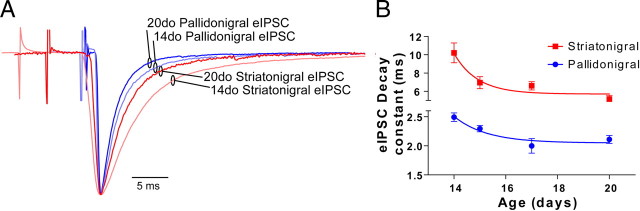
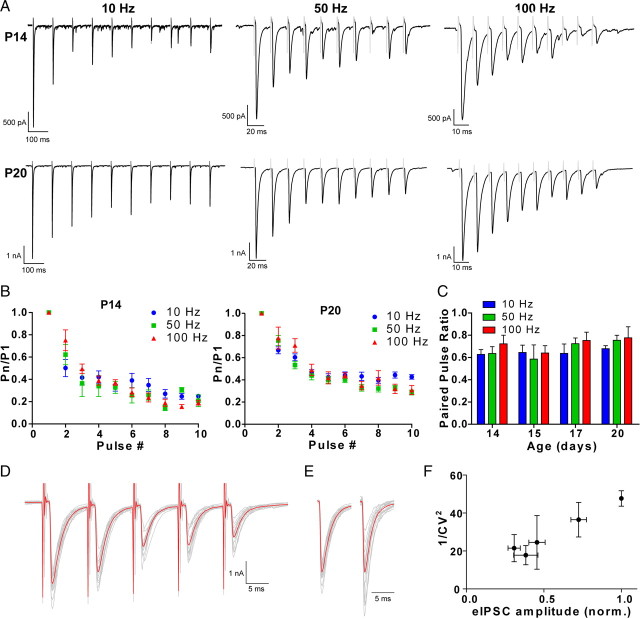
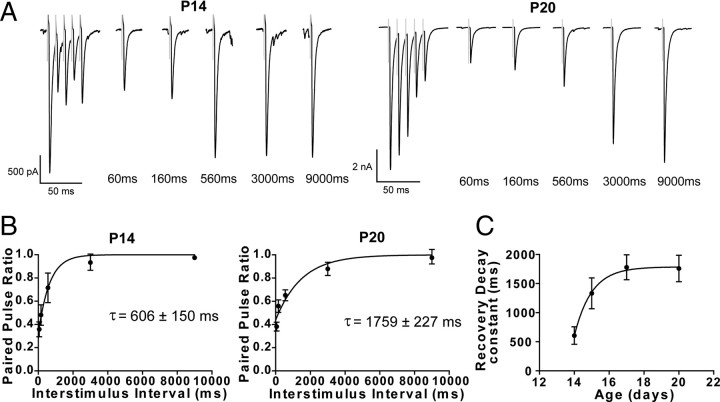
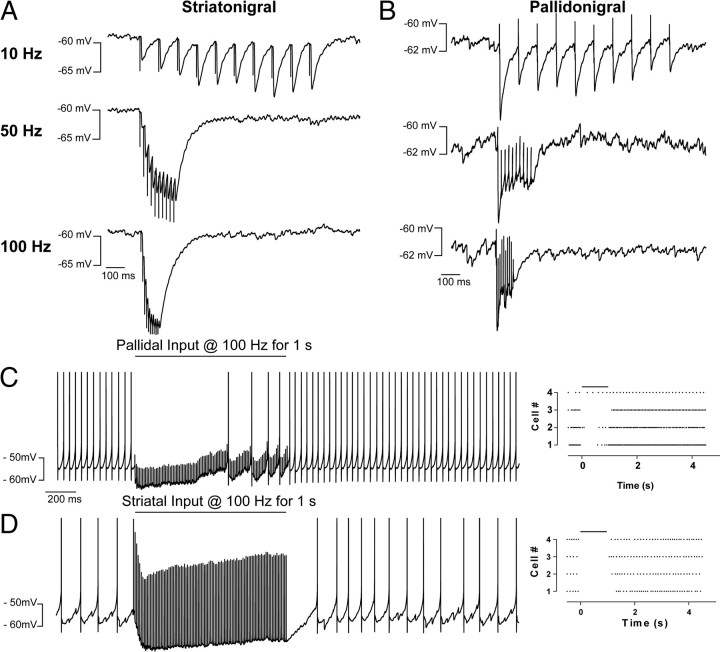
Inhibitory projections from the striatum and globus pallidus converge onto GABAergic projection neurons of the substantia nigra pars reticulata (SNr). Based on existing structural and functional evidence, these pathways are likely to differentially regulate the firing of SNr neurons. We sought to investigate the functional differences in inhibitory striatonigral and pallidonigral traffic using whole-cell voltage clamp in brain slices with these pathways preserved. We found that striatonigral IPSCs exhibited a high degree of paired-pulse facilitation. We tracked this facilitation over development and found the facilitation as the animal aged, but stabilized by postnatal day 17 (P17), with a paired pulse ratio of 2. We also found that the recovery from facilitation accelerated over development, again, reaching a stable phenotype by P17. In contrast, pallidonigral synapses show paired-pulse depression, and this depression could be solely explained by presynaptic changes. The mean paired-pulse ratio of 0.67 did not change over development, but the recovery from depression slowed over development. Pallidonigral IPSCs were significantly faster than striatonigral IPSCs when measured at the soma. Finally, under current clamp, prolonged bursts of striatal IPSPs were able to consistently silence the pacemaker activity of nigral neurons, whereas pallidal inputs depressed, allowing nigral neurons to reinstate firing. These findings highlight the importance of differential dynamics of neurotransmitter release in regulating the circuit behavior of the basal ganglia.
Figures



References
-
- Araki M, McGeer PL, McGeer EG. Striatonigral and pallidonigral pathways studied by a combination of retrograde horseradish peroxidase tracing and a pharmacohistochemical method for gamma-aminobutyric acid transaminase. Brain Res. 1985;331:17–24. - PubMed
-
- Barnes TD, Kubota Y, Hu D, Jin DZ, Graybiel AM. Activity of striatal neurons reflects dynamic encoding and recoding of procedural memories. Nature. 2005;437:1158–1161. - PubMed
-
- Beurrier C, Ben-Ari Y, Hammond C. Preservation of the direct and indirect pathways in an in vitro preparation of the mouse basal ganglia. Neuroscience. 2006;140:77–86. - PubMed
-
- Chang HT, Wilson CJ, Kitai ST. Single neostriatal efferent axons in the globus pallidus: a light and electron microscopic study. Science. 1981;213:915–918. - PubMed
-
- Chevalier G, Deniau JM. Disinhibition as a basic process in the expression of striatal functions. Trends Neurosci. 1990;13:277–280. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous