

Mammalian circadian clock and metabolism - the epigenetic link
- PMID: 21048160
- PMCID: PMC2972271
- DOI: 10.1242/jcs.051649
Mammalian circadian clock and metabolism - the epigenetic link
Abstract
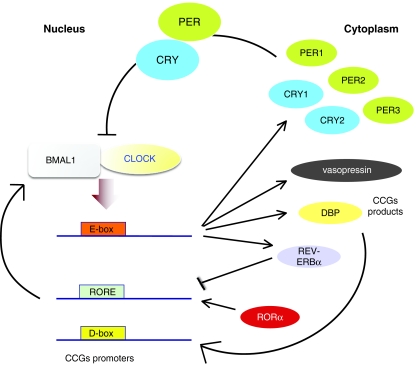
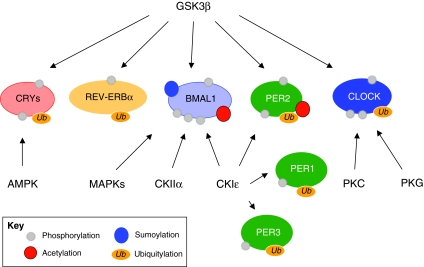
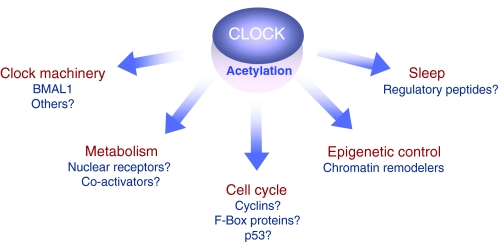
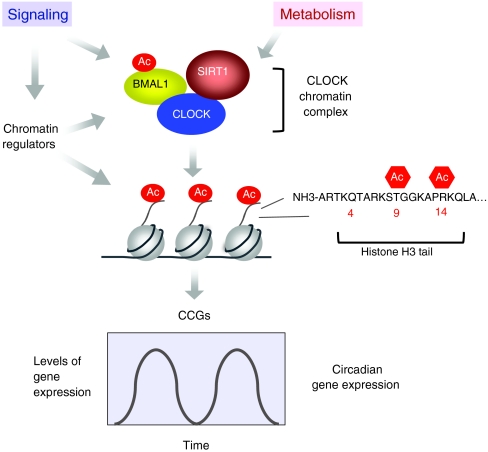
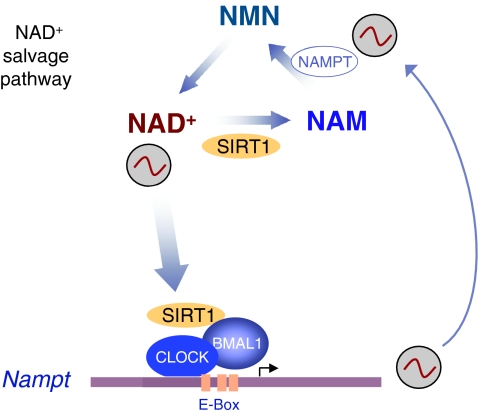
Circadian rhythms regulate a wide variety of physiological and metabolic processes. The clock machinery comprises complex transcriptional-translational feedback loops that, through the action of specific transcription factors, modulate the expression of as many as 10% of cellular transcripts. This marked change in gene expression necessarily implicates a global regulation of chromatin remodeling. Indeed, various descriptive studies have indicated that histone modifications occur at promoters of clock-controlled genes (CCGs) in a circadian manner. The finding that CLOCK, a transcription factor crucial for circadian function, has intrinsic histone acetyl transferase (HAT) activity has paved the way to unraveling the molecular mechanisms that govern circadian chromatin remodeling. A search for the histone deacetylase (HDAC) that counterbalances CLOCK activity revealed that SIRT1, a nicotinamide adenin dinucleotide (NAD(+))-dependent HDAC, functions in a circadian manner. Importantly, SIRT1 is a regulator of aging, inflammation and metabolism. As many transcripts that oscillate in mammalian peripheral tissues encode proteins that have central roles in metabolic processes, these findings establish a functional and molecular link between energy balance, chromatin remodeling and circadian physiology. Here we review recent studies that support the existence of this link and discuss their implications for understanding mammalian physiology and pathology.
Figures
References
-
- Akashi M., Takumi T. (2005). The orphan nuclear receptor RORalpha regulates circadian transcription of the mammalian core-clock Bmal1. Nat. Struct. Mol. Biol. 12, 441-448 - PubMed
-
- Akhtar R. A., Reddy A. B., Maywood E. S., Clayton J. D., King V. M., Smith A. G., Gant T. W., Hastings M. H., Kyriacou C. P. (2002). Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr. Biol. 12, 540-550 - PubMed
-
- Akten B., Jauch E., Genova G. K., Kim E. Y., Edery I., Raabe T., Jackson F. R. (2003). A role for CK2 in the Drosophila circadian oscillator. Nat. Neurosci. 6, 251-257 - PubMed
-
- Ando H., Yanagihara H., Hayashi Y., Obi Y., Tsuruoka S., Takamura T., Kaneko S., Fujimura A. (2005). Rhythmic messenger ribonucleic acid expression of clock genes and adipocytokines in mouse visceral adipose tissue. Endocrinology 146, 5631-5636 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
