

Ultrastructural characterization of noradrenergic axons and Beta-adrenergic receptors in the lateral nucleus of the amygdala
- PMID: 21048893
- PMCID: PMC2967335
- DOI: 10.3389/fnbeh.2010.00162
Ultrastructural characterization of noradrenergic axons and Beta-adrenergic receptors in the lateral nucleus of the amygdala
Abstract
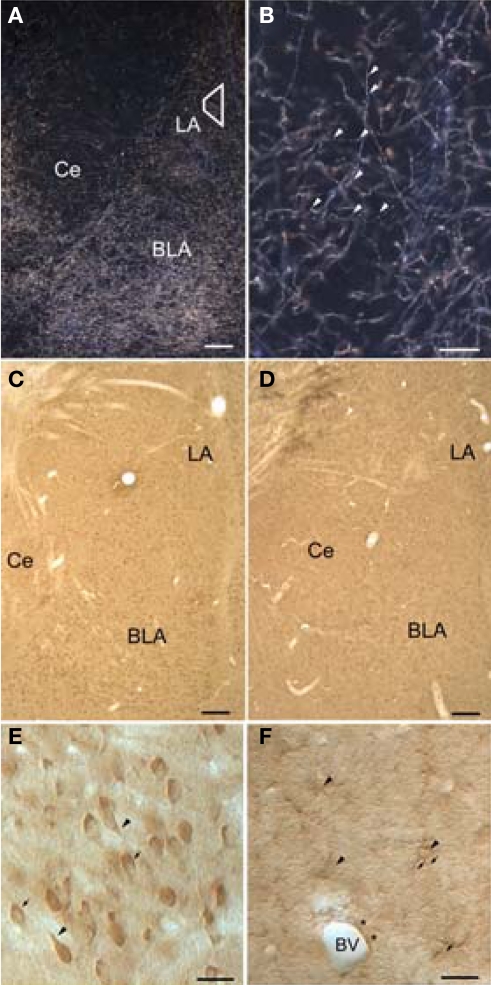
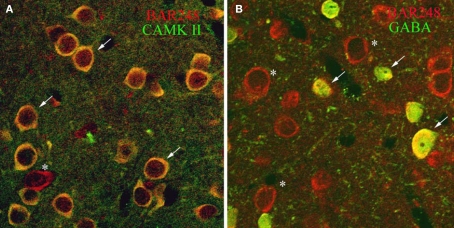
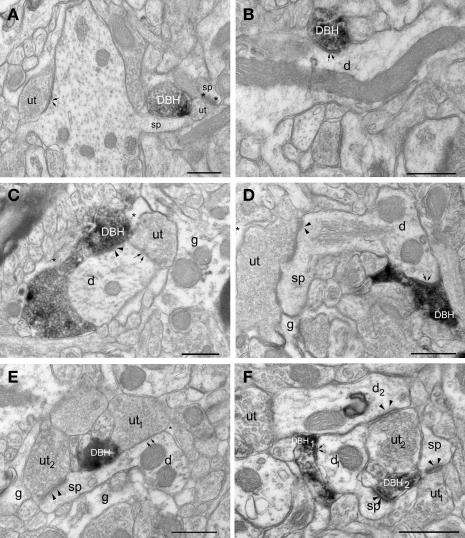
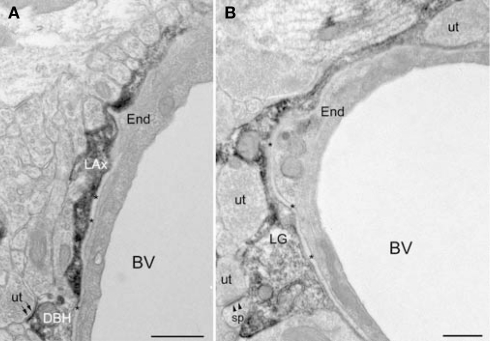
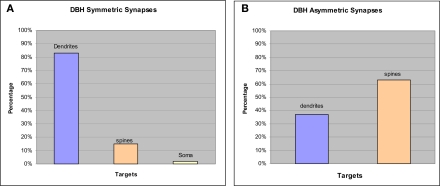
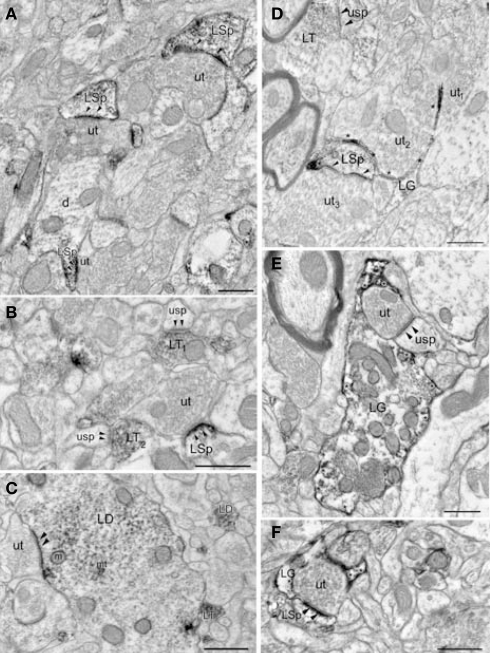
Norepinephrine (NE) is thought to play a key role in fear and anxiety, but its role in amygdala-dependent Pavlovian fear conditioning, a major model for understanding the neural basis of fear, is poorly understood. The lateral nucleus of the amygdala (LA) is a critical brain region for fear learning and regulating the effects of stress on memory. To understand better the cellular mechanisms of NE and its adrenergic receptors in the LA, we used antibodies directed against dopamine beta-hydroxylase (DβH), the synthetic enzyme for NE, or against two different isoforms of the beta-adrenergic receptors (βARs), one that predominately recognizes neurons (βAR 248) and the other astrocytes (βAR 404), to characterize the microenvironments of DβH and βAR. By electron microscopy, most DβH terminals did not make synapses, but when they did, they formed both asymmetric and symmetric synapses. By light microscopy, βARs were present in both neurons and astrocytes. Confocal microscopy revealed that both excitatory and inhibitory neurons express βAR248. By electron microscopy, βAR 248 was present in neuronal cell bodies, dendritic shafts and spines, and some axon terminals and astrocytes. When in dendrites and spines, βAR 248 was frequently concentrated along plasma membranes and at post-synaptic densities of asymmetric (excitatory) synapses. βAR 404 was expressed predominately in astrocytic cell bodies and processes. These astrocytic processes were frequently interposed between unlabeled terminals or ensheathed asymmetric synapses. Our findings provide a morphological basis for understanding ways in which NE may modulate transmission by acting via synaptic or non-synaptic mechanisms in the LA.
Keywords: DBH; astrocyte; electron microscopy; fear; lateral amygdala; norepinephrine; synapse; volume transmission.
Figures
Similar articles
-
Synapses lacking astrocyte appear in the amygdala during consolidation of Pavlovian threat conditioning.J Comp Neurol. 2014 Jun 15;522(9):2152-63. doi: 10.1002/cne.23523. J Comp Neurol. 2014. PMID: 24338694 Free PMC article.
-
Noradrenergic innervation of pyramidal cells in the rat basolateral amygdala.Neuroscience. 2013 Jan 3;228:395-408. doi: 10.1016/j.neuroscience.2012.10.035. Epub 2012 Oct 26. Neuroscience. 2013. PMID: 23103792 Free PMC article.
-
Synapses on GABAergic neurons in the basolateral nucleus of the rat amygdala: double-labeling immunoelectron microscopy.Synapse. 2002 Jan;43(1):42-50. doi: 10.1002/syn.10017. Synapse. 2002. PMID: 11746732
-
Beta-adrenergic receptors in the lateral nucleus of the amygdala contribute to the acquisition but not the consolidation of auditory fear conditioning.Front Behav Neurosci. 2010 Oct 26;4:154. doi: 10.3389/fnbeh.2010.00154. eCollection 2010. Front Behav Neurosci. 2010. PMID: 21152344 Free PMC article.
-
beta-adrenergic receptors primarily are located on the dendrites of granule cells and interneurons but also are found on astrocytes and a few presynaptic profiles in the rat dentate gyrus.Synapse. 2000 Jun 1;36(3):178-93. doi: 10.1002/(SICI)1098-2396(20000601)36:3<178::AID-SYN3>3.0.CO;2-6. Synapse. 2000. PMID: 10819898
Cited by
-
Synaptic competition in the lateral amygdala and the stimulus specificity of conditioned fear: a biophysical modeling study.Brain Struct Funct. 2016 May;221(4):2163-82. doi: 10.1007/s00429-015-1037-4. Epub 2015 Apr 10. Brain Struct Funct. 2016. PMID: 25859631 Free PMC article.
-
Norepinephrine, beyond the Synapse: Coordinating Epigenetic Codes for Memory.Int J Mol Sci. 2022 Aug 31;23(17):9916. doi: 10.3390/ijms23179916. Int J Mol Sci. 2022. PMID: 36077313 Free PMC article. Review.
-
Norepinephrine Signals Through Astrocytes To Modulate Synapses.bioRxiv [Preprint]. 2024 May 22:2024.05.21.595135. doi: 10.1101/2024.05.21.595135. bioRxiv. 2024. Update in: Science. 2025 May 15;388(6748):776-783. doi: 10.1126/science.adq5480. PMID: 38826209 Free PMC article. Updated. Preprint.
-
Molecular mechanisms of fear learning and memory.Cell. 2011 Oct 28;147(3):509-24. doi: 10.1016/j.cell.2011.10.009. Cell. 2011. PMID: 22036561 Free PMC article. Review.
-
Kinase and Phosphatase Engagement Is Dissociated Between Memory Formation and Extinction.Front Mol Neurosci. 2019 Feb 20;12:38. doi: 10.3389/fnmol.2019.00038. eCollection 2019. Front Mol Neurosci. 2019. PMID: 30842725 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous