

Androgen action via testicular arteriole smooth muscle cells is important for Leydig cell function, vasomotion and testicular fluid dynamics
- PMID: 21049031
- PMCID: PMC2964321
- DOI: 10.1371/journal.pone.0013632
Androgen action via testicular arteriole smooth muscle cells is important for Leydig cell function, vasomotion and testicular fluid dynamics
Abstract
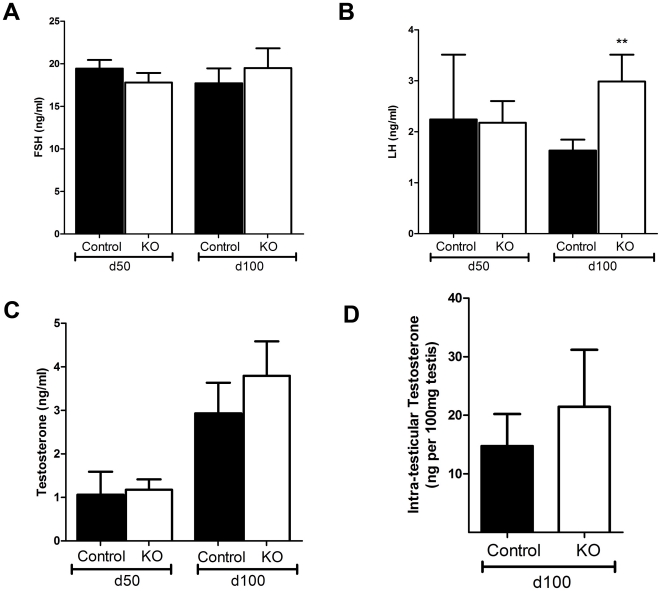
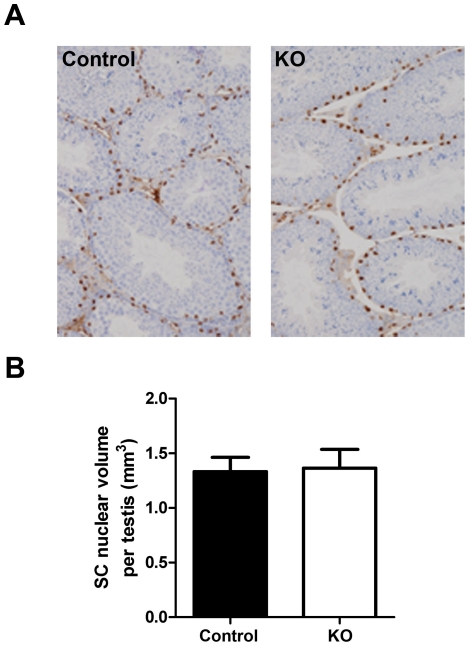
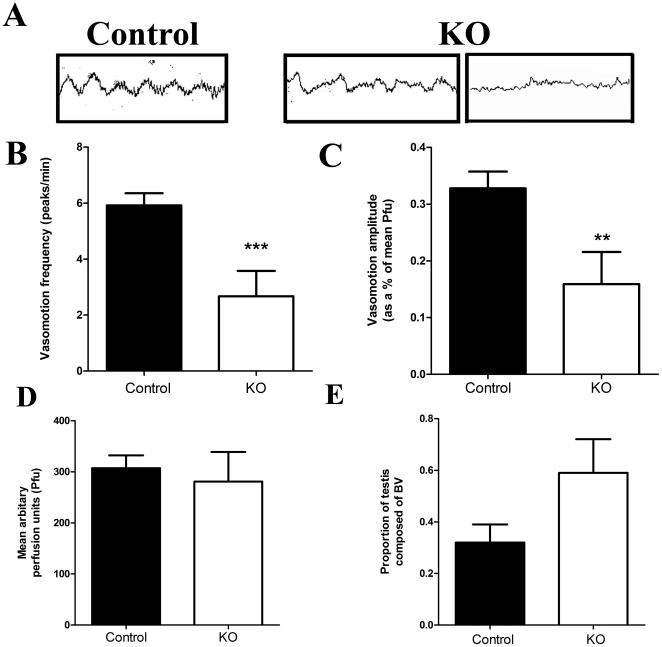
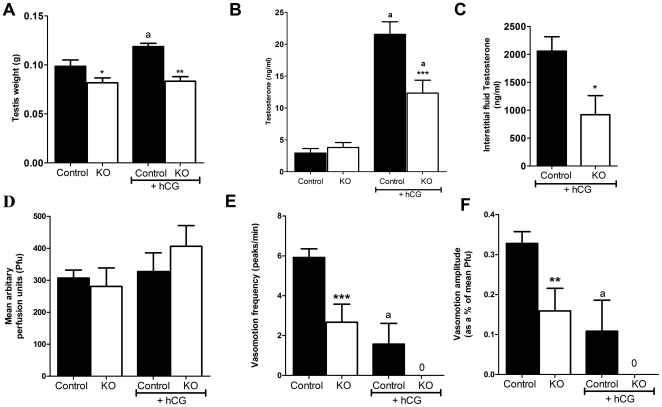
Regulation of blood flow through the testicular microvasculature by vasomotion is thought to be important for normal testis function as it regulates interstitial fluid (IF) dynamics which is an important intra-testicular transport medium. Androgens control vasomotion, but how they exert these effects remains unclear. One possibility is by signalling via androgen receptors (AR) expressed in testicular arteriole smooth muscle cells. To investigate this and determine the overall importance of this mechanism in testis function, we generated a blood vessel smooth muscle cell-specific AR knockout mouse (SMARKO). Gross reproductive development was normal in SMARKO mice but testis weight was reduced in adulthood compared to control littermates; this reduction was not due to any changes in germ cell volume or to deficits in testosterone, LH or FSH concentrations and did not cause infertility. However, seminiferous tubule lumen volume was reduced in adult SMARKO males while interstitial volume was increased, perhaps indicating altered fluid dynamics; this was associated with compensated Leydig cell failure. Vasomotion was impaired in adult SMARKO males, though overall testis blood flow was normal and there was an increase in the overall blood vessel volume per testis in adult SMARKOs. In conclusion, these results indicate that ablating arteriole smooth muscle AR does not grossly alter spermatogenesis or affect male fertility but does subtly impair Leydig cell function and testicular fluid exchange, possibly by locally regulating microvascular blood flow within the testis.
Conflict of interest statement
Figures
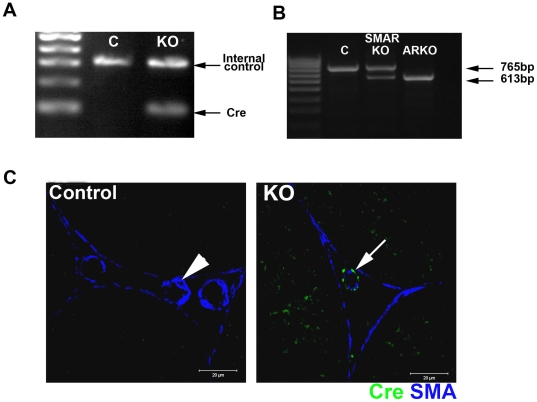
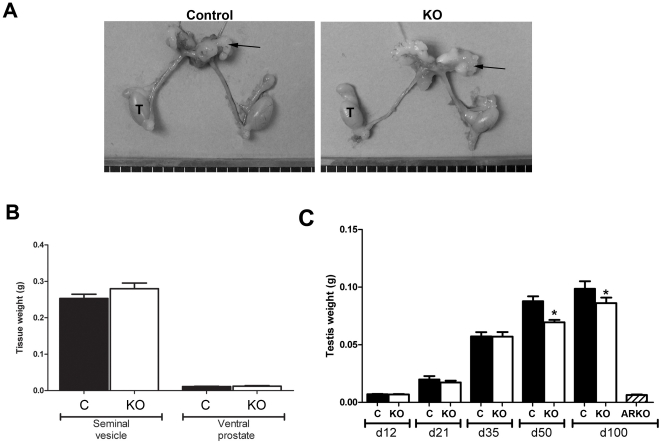
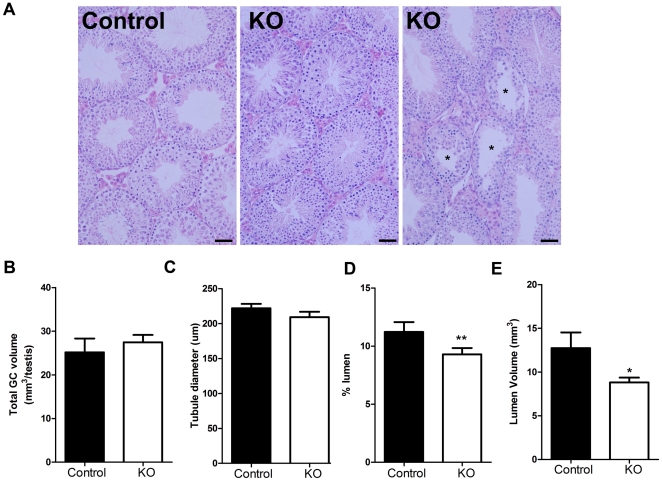
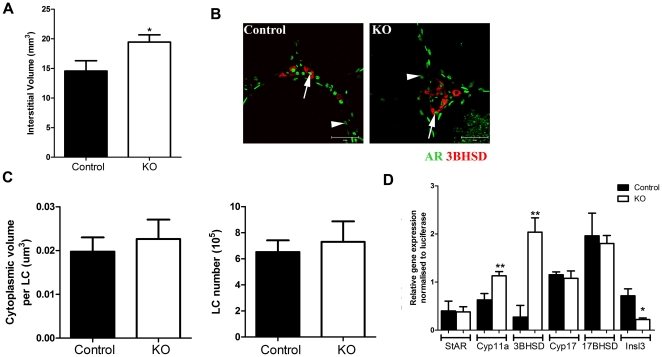
References
-
- George FW, Wilson J. Gonads and ducts in mammals. In: Knobil E, Neill, JD, editors. The Physiology of Reproduction. 2nd ed. New York: Raven Press; 1994. pp. 3–27.
-
- McLachlan RI, O'Donnell L, Meachem SJ, Stanton PG, de Kretser DM, et al. Identification of specific sites of hormonal regulation in spermatogenesis in rats, monkeys, and man. Recent Prog Horm Res. 2002;57:149–179. - PubMed
-
- Sharpe RM. The Physiology of Reproduction. In: Knobil EaN JD, editor. New York: Raven; 1994. pp. 1363–2434.
-
- Quigley CA, De Bellis A, Marschke KB, el-Awady MK, Wilson EM, et al. Androgen receptor defects: historical, clinical, and molecular perspectives. Endocr Rev. 1995;16:271–321. - PubMed
-
- Bremner WJ, Millar MR, Sharpe RM, Saunders PT. Immunohistochemical localization of androgen receptors in the rat testis: evidence for stage-dependent expression and regulation by androgens. Endocrinology. 1994;135:1227–1234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_U127685841/MRC_/Medical Research Council/United Kingdom
- MC_U127685844/MRC_/Medical Research Council/United Kingdom
- MC_U127684422/MRC_/Medical Research Council/United Kingdom
- U.1276.00.002.00003.01 (85841)/MRC_/Medical Research Council/United Kingdom
- WBS U.1276.00.002.0003.01/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
