

Hearing it again and again: on-line subcortical plasticity in humans
- PMID: 21049035
- PMCID: PMC2964325
- DOI: 10.1371/journal.pone.0013645
Hearing it again and again: on-line subcortical plasticity in humans
Abstract
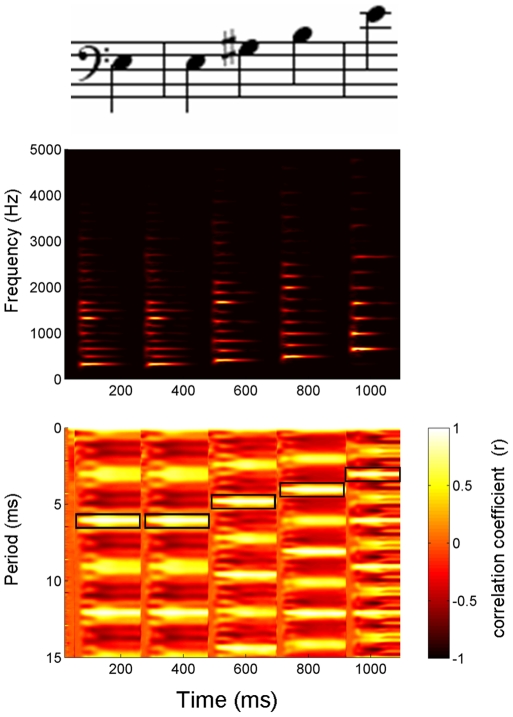
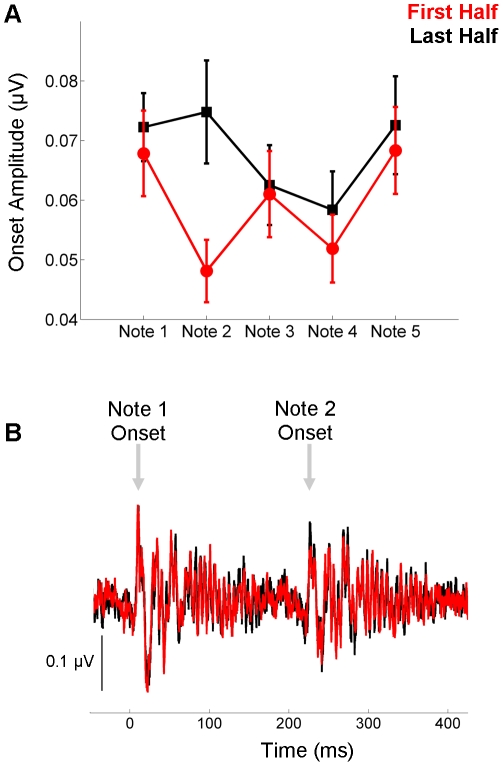
Background: Human brainstem activity is sensitive to local sound statistics, as reflected in an enhanced response in repetitive compared to pseudo-random stimulus conditions [1]. Here we probed the short-term time course of this enhancement using a paradigm that assessed how the local sound statistics (i.e., repetition within a five-note melody) interact with more global statistics (i.e., repetition of the melody).
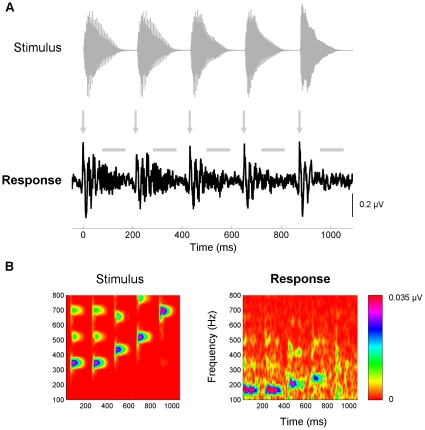
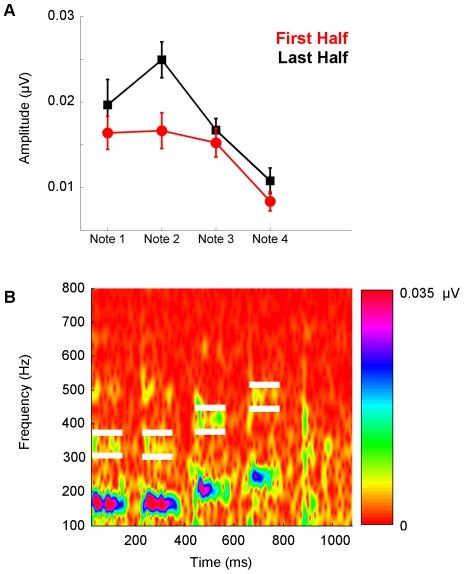
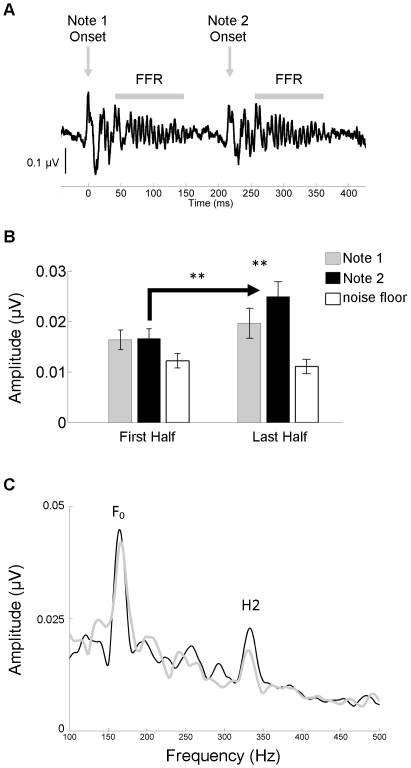
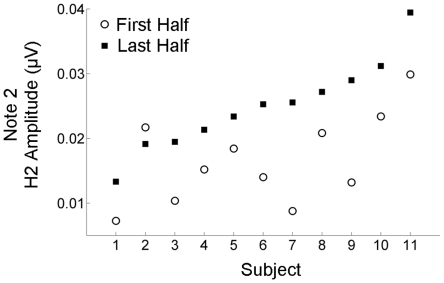
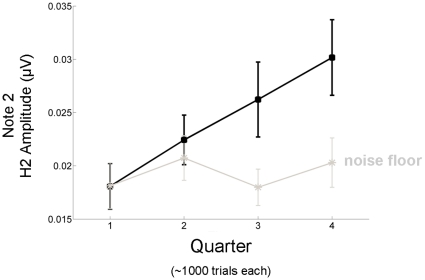
Methodology/principal findings: To test the hypothesis that subcortical repetition enhancement builds over time, we recorded auditory brainstem responses in young adults to a five-note melody containing a repeated note, and monitored how the response changed over the course of 1.5 hrs. By comparing response amplitudes over time, we found a robust time-dependent enhancement to the locally repeating note that was superimposed on a weaker enhancement of the globally repeating pattern.
Conclusions/significance: We provide the first demonstration of on-line subcortical plasticity in humans. This complements previous findings that experience-dependent subcortical plasticity can occur on a number of time scales, including life-long experiences with music and language, and short-term auditory training. Our results suggest that the incoming stimulus stream is constantly being monitored, even when the stimulus is physically invariant and attention is directed elsewhere, to augment the neural response to the most statistically salient features of the ongoing stimulus stream. These real-time transformations, which may subserve humans' strong disposition for grouping auditory objects, likely reflect a mix of local processes and corticofugal modulation arising from statistical regularities and the influences of expectation. Our results contribute to our understanding of the biological basis of statistical learning and initiate a new investigational approach relating to the time-course of subcortical plasticity. Although the reported time-dependent enhancements are believed to reflect universal neurophysiological processes, future experiments utilizing a larger array of stimuli are needed to establish the generalizability of our findings.
Conflict of interest statement
Figures
References
-
- Large E, Jones M. The dynamics of attending: How people track time-varying events. Psychol Rev. 1999;106:119–159.
-
- Saffran JR. Musical learning and language development. Ann N Y Acad Sci. 2003;999:397–401. - PubMed
-
- Winkler I, Denham SL, Nelken I. Modeling the auditory scene: predictive regularity representations and perceptual objects. Trends Cogn Sci. 2009;13:532–540. - PubMed
-
- Drake C, Bertrand D. The quest for universals in temporal processing in music. Ann N Y Acad Sci. 2001;930:17–27. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
