

Direct involvement of the TEN domain at the active site of human telomerase
- PMID: 21051362
- PMCID: PMC3061064
- DOI: 10.1093/nar/gkq1083
Direct involvement of the TEN domain at the active site of human telomerase
Abstract
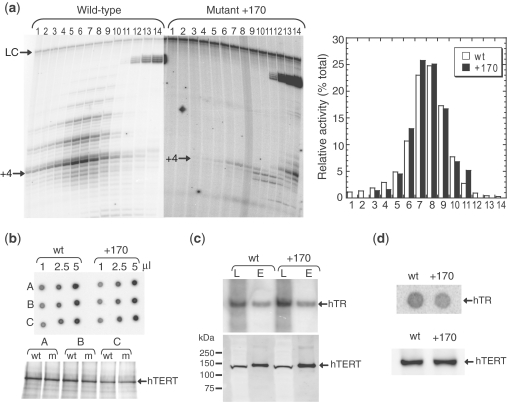
Telomerase is a ribonucleoprotein that adds DNA to the ends of chromosomes. The catalytic protein subunit of telomerase (TERT) contains an N-terminal domain (TEN) that is important for activity and processivity. Here we describe a mutation in the TEN domain of human TERT that results in a greatly increased primer K(d), supporting a role for the TEN domain in DNA affinity. Measurement of enzyme kinetic parameters has revealed that this mutant enzyme is also defective in dNTP polymerization, particularly while copying position 51 of the RNA template. The catalytic defect is independent of the presence of binding interactions at the 5'-region of the DNA primer, and is not a defect in translocation rate. These data suggest that the TEN domain is involved in conformational changes required to position the 3'-end of the primer in the active site during nucleotide addition, a function which is distinct from the role of the TEN domain in providing DNA binding affinity.
Figures






Similar articles
-
A mutation in the catalytic subunit of yeast telomerase alters primer-template alignment while promoting processivity and protein-DNA binding.J Cell Sci. 2011 Dec 15;124(Pt 24):4241-52. doi: 10.1242/jcs.090761. Epub 2011 Dec 22. J Cell Sci. 2011. PMID: 22193961 Free PMC article.
-
The telomerase essential N-terminal domain promotes DNA synthesis by stabilizing short RNA-DNA hybrids.Nucleic Acids Res. 2015 Jun 23;43(11):5537-49. doi: 10.1093/nar/gkv406. Epub 2015 May 4. Nucleic Acids Res. 2015. PMID: 25940626 Free PMC article.
-
Multiple DNA-binding sites in Tetrahymena telomerase.Nucleic Acids Res. 2008 Mar;36(4):1260-72. doi: 10.1093/nar/gkm866. Epub 2008 Jan 3. Nucleic Acids Res. 2008. PMID: 18174223 Free PMC article.
-
Structure and function of telomerase RNA.Curr Opin Struct Biol. 2006 Jun;16(3):307-18. doi: 10.1016/j.sbi.2006.05.005. Epub 2006 May 18. Curr Opin Struct Biol. 2006. PMID: 16713250 Review.
-
Evolutionary perspectives of telomerase RNA structure and function.RNA Biol. 2016 Aug 2;13(8):720-32. doi: 10.1080/15476286.2016.1205768. Epub 2016 Jun 30. RNA Biol. 2016. PMID: 27359343 Free PMC article. Review.
Cited by
-
Telomerase recruitment requires both TCAB1 and Cajal bodies independently.Mol Cell Biol. 2012 Jul;32(13):2384-95. doi: 10.1128/MCB.00379-12. Epub 2012 Apr 30. Mol Cell Biol. 2012. PMID: 22547674 Free PMC article.
-
Structure of Telomerase with Telomeric DNA.Cell. 2018 May 17;173(5):1179-1190.e13. doi: 10.1016/j.cell.2018.04.038. Cell. 2018. PMID: 29775593 Free PMC article.
-
A mutation in the catalytic subunit of yeast telomerase alters primer-template alignment while promoting processivity and protein-DNA binding.J Cell Sci. 2011 Dec 15;124(Pt 24):4241-52. doi: 10.1242/jcs.090761. Epub 2011 Dec 22. J Cell Sci. 2011. PMID: 22193961 Free PMC article.
-
Single-Molecule Studies of Telomeres and Telomerase.Annu Rev Biophys. 2017 May 22;46:357-377. doi: 10.1146/annurev-biophys-062215-011256. Epub 2017 Mar 22. Annu Rev Biophys. 2017. PMID: 28375735 Free PMC article. Review.
-
Single-molecule imaging of telomerase reverse transcriptase in human telomerase holoenzyme and minimal RNP complexes.Elife. 2015 Oct 12;4:e08363. doi: 10.7554/eLife.08363. Elife. 2015. PMID: 26457608 Free PMC article.
References
-
- McEachern MJ, Krauskopf A, Blackburn EH. Telomeres and their control. Annu. Rev. Genet. 2000;34:331–358. - PubMed
-
- Greider CW, Blackburn EH. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 1985;43:405–413. - PubMed
-
- Shay JW, Bacchetti S. A survey of telomerase activity in human cancer. Eur. J. Cancer. 1997;33:787–791. - PubMed
-
- Hahn WC, Stewart SA, Brooks MW, York SG, Eaton E, Kurachi A, Beijersbergen RL, Knoll JH, Meyerson M, Weinberg RA. Inhibition of telomerase limits the growth of human cancer cells. Nat. Med. 1999;5:1164–1170. - PubMed
