

Review
doi: 10.1104/pp.110.163360.
Epub 2010 Nov 4.
Update on mechanisms of plant cell wall biosynthesis: how plants make cellulose and other (1->4)-β-D-glycans
Affiliations
- PMID: 21051553
- PMCID: PMC3075763
- DOI: 10.1104/pp.110.163360
Item in Clipboard
Review
Update on mechanisms of plant cell wall biosynthesis: how plants make cellulose and other (1->4)-β-D-glycans
Plant Physiol.
2011 Jan.
No abstract available
Figures

The CesA/Csl gene superfamily. A, Of the 10 Arabidopsis CesA genes, at least three are coexpressed during primary wall formation, and mutations in each of them, AtCesA1 (RSW1; At4g32410), AtCesA6 (PRC1; At5g64740), and AtCesA3 (CEV1 and ELI1; At5g05170), result in cellulose deficiencies, indicating that each is essential for cellulose synthesis. The irx mutants AtCesA8 (IRX1; At4g18780), AtCesA7 (IRX3; At5g17420), and AtCesA4 (IRX5; At5g44030) are deficient in cellulose synthesis specifically in secondary walls. Seven additional subgroups were identified that are the likely synthases for noncellulosic polysaccharides with backbones of (1→4)-β-d -glycans. Whereas the CesA genes of Arabidopsis, rice, and maize appear to be orthologous, the Csl genes are divergent between dicots and grasses, species that make two distinct kinds of walls. From mutants and heterologous expression studies, members of the CslA group encode the synthases of (gluco)mannans, members of the CslC group are likely to encode the glucan backbone of xyloglucans, and the rice- and maize-only members of CslH and CslF encode the synthases of the mixed-linkage (1→3),(1→4)-β-d -glucans found only in grasses (after Penning et al., 2009). B, Domain model and class-specific regions (CSRs) for three CesAs known to function in primary cell wall cellulose synthesis. Two ZnF domains (in yellow) are found in the N terminus before the first membrane-spanning domain (in blue). Eight transmembrane helices, two upstream and six downstream of the catalytic domain, are predicted to interact to form a channel through which a single β-glucan chain is secreted to the cell wall. The large central catalytic domain contains four highly conserved “U motifs” of D, DxD, D, and QxxRW, important for substrate binding and catalysis. Once thought to be a hypervariable region (Pear et al., 1996), the class-specific regions are conserved among orthologs of the same subclade and vary in the number of upstream conserved Cys residues, the number of consecutive basic amino acids, Lys and Arg, and the number of consecutive acidic amino acids, Asp and Glu, downstream from the basic residues (after Carpita and Vergara, 1998; Vergara and Carpita, 2001).

Particle rosette structures associated with cellulose synthesis in angiosperms. A, Freeze-etch images of the P-face of the plasma membrane showing clusters of rosettes associated with the developing of secondary wall spiral thickenings of a Lepidium tracheary element (from Herth, 1985). The inset shows the 6-fold symmetry of a single particle rosette from a Zinnia tracheary element developing in vitro (from C. Haigler, unpublished data, as seen in Delmer, 1999). A substructure can be observed in each of the particles. In these freeze-etch images, only the membrane-spanning domains and extracellular loops of the CesA proteins can be observed. B, Cytoplasmic structure (circled) underlying the rosettes in plasma membrane footprints (from Bowling and Brown, 2008). These structures always are at the terminus of a microfibril (arrow). Bar = 200 nm. C, A Markham rotational analysis of one of these shows the reinforcement of hexagonal shape with 60° rotational steps. All other angles of rotation cancel to circular (Bowling and Brown, 2008).

Models for cellulose synthase and the steric problem of making a (1→4)-β-d -glycosyl linkage. A, The first model of conformation for a single CesA protein subunit was proposed by Delmer (1999). Each CesA subunit must interact with other such subunits to form the synthase complex. The ZnFs, plant-specific conserved region (P-CR), and class-specific region (CSR) are potential interaction sites (figure modified from Delmer, 1999). B, The steric problem of synthesis is illustrated in top view and end view. Addition of a single glycosyl residue in a (1→4)-β-d linkage without rotation of the end of the chain or the active site of the synthase would result in movement of the O-4 several angstroms. C, The conceptual solution to the steric problem is a catalytic dimer of simultaneous glycosyl transfer to form a cellobiosyl residue to the O-4 position of the terminal glucosyl residue of the chain. Synthesis of an even number of units always maintains the acceptor position as the O-4 position as the chain is extruded. In the catalytic dimer model, if one of the sites is damaged, then the point of attachment becomes the O-3 position, which would maintain the point of attachment as the O-3, hence producing callose thereafter (after Buckeridge et al., 1999, 2001).

Models of HA synthases. A, Class I synthases. B, Class II synthases (after Weigel and DeAngelis, 2007).
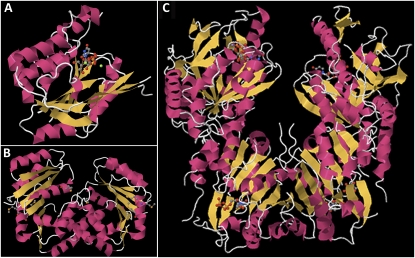
Crystal structures of type 2 glycosyl transferases. A, The SpsA synthase crystallizes as a monomer with a single binding site for UDP (Charnock and Davies, 1999; http://www.pdb.org/pdb/explore/explore.do?structureId=1QGS ). B, The B. fragilis SpsA homologous polypeptide crystallizes as a dimer (http://www.pdb.org/pdb/explore/explore.do?structureId=3BCV ), each with a single binding domain for UDP. C, Each member of the crystal dimer E. coli chondroitin polymerase has two UDP-GlcA- or UDP-binding domains (http://www.pdb.org/pdb/explore/explore.do?structureId=2Z86 ).

A catalytic dimer hypothesis for cellulose synthase. A, A catalytic dimer model of two CesAs to form a complex that synthesizes a single (1→4)-β-d -glucan chain. Homodimerization or heterodimerization of CesAs gives mirrored active sites that generate cellobiosyl units, which are then attached to the nonreducing end of the extruded glucan chain. Dimerization also results in a channel composed of 16 membrane-spanning domains, equivalent to that of callose synthase and consistent with eukaryotic monosaccharide transporters. B, Dimerization results in two ZnF domains that are now able to couple two neighbors instead of just one. C, Six such complexes interact to constitute one particle of the six-particle rosette.
Similar articles
-
Cellulose synthesis: a complex complex.Curr Opin Plant Biol. 2008 Jun;11(3):252-7. doi: 10.1016/j.pbi.2008.03.007. Epub 2008 May 14. Curr Opin Plant Biol. 2008. PMID: 18485800 Review.
-
Phosphorylation of cellulose synthases in plant responses to environmental changes.Int J Biol Macromol. 2025 Mar;292:139313. doi: 10.1016/j.ijbiomac.2024.139313. Epub 2024 Dec 29. Int J Biol Macromol. 2025. PMID: 39740718 Review.
-
Biosynthesis of cellulose.Adv Carbohydr Chem Biochem. 1983;41:105-53. doi: 10.1016/s0065-2318(08)60057-8. Adv Carbohydr Chem Biochem. 1983. PMID: 6227209 Review. No abstract available.
-
Cellulose microfibrils in plants: biosynthesis, deposition, and integration into the cell wall.Int Rev Cytol. 2000;199:161-99. doi: 10.1016/s0074-7696(00)99004-1. Int Rev Cytol. 2000. PMID: 10874579 Review.
-
Cellulose metabolism in plants.Int Rev Cytol. 2005;247:1-34. doi: 10.1016/S0074-7696(05)47001-1. Int Rev Cytol. 2005. PMID: 16344110 Review.
Cited by
-
Purification and characterization of a soluble β-1,4-glucan from bean (Phaseolus vulgaris L.)-cultured cells dehabituated to dichlobenil.Planta. 2013 Jun;237(6):1475-82. doi: 10.1007/s00425-013-1861-9. Epub 2013 Feb 28. Planta. 2013. PMID: 23455460
-
Mannan synthase activity in the CSLD family.Plant Signal Behav. 2011 Oct;6(10):1620-3. doi: 10.4161/psb.6.10.17989. Epub 2011 Oct 1. Plant Signal Behav. 2011. PMID: 21904114 Free PMC article.
-
Insights into the structure and function of membrane-integrated processive glycosyltransferases.Curr Opin Struct Biol. 2015 Oct;34:78-86. doi: 10.1016/j.sbi.2015.07.008. Epub 2015 Sep 2. Curr Opin Struct Biol. 2015. PMID: 26342143 Free PMC article. Review.
-
The reference genome of Miscanthus floridulus illuminates the evolution of Saccharinae.Nat Plants. 2021 May;7(5):608-618. doi: 10.1038/s41477-021-00908-y. Epub 2021 May 6. Nat Plants. 2021. PMID: 33958777 Free PMC article.
-
Cellulose microfibril crystallinity is reduced by mutating C-terminal transmembrane region residues CESA1A903V and CESA3T942I of cellulose synthase.Proc Natl Acad Sci U S A. 2012 Mar 13;109(11):4098-103. doi: 10.1073/pnas.1200352109. Epub 2012 Feb 28. Proc Natl Acad Sci U S A. 2012. PMID: 22375033 Free PMC article.
References
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815 - PubMed
-
- Arioli T, Peng LC, Betzner AS, Burn J, Wittke W, Herth W, Camilleri C, Höfte H, Plazinski J, Birch R, et al. (1998) Molecular analysis of cellulose biosynthesis in Arabidopsis. Science 279: 717–720 - PubMed
-
- Atanassov II, Pittman JK, Turner SR. (2009) Elucidating the mechanisms of assembly and subunit interaction of the cellulose synthase complex of Arabidopsis secondary cell walls. J Biol Chem 284: 3833–3841 - PubMed
-
- Bacic A, Delmer DP. (1981) Stimulation of membrane-associated polysaccharide synthetases by a membrane potential in developing cotton fibers. Planta 152: 346–351 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
