

Protease inhibitor-resistant hepatitis C virus mutants with reduced fitness from impaired production of infectious virus
- PMID: 21056040
- PMCID: PMC3155954
- DOI: 10.1053/j.gastro.2010.10.056
Protease inhibitor-resistant hepatitis C virus mutants with reduced fitness from impaired production of infectious virus
Abstract
Background & aims: Several small molecule inhibitors of the hepatitis C virus (HCV) nonstructural protein (NS) 3/4A protease have advanced successfully to clinical trials. However, the selection of drug-resistant mutants is a significant issue with protease inhibitors (PIs). A variety of amino acid substitutions in the protease domain of NS3 can lead to PI resistance. Many of these significantly impair the replication fitness of HCV RNA replicons. However, it is not known whether these mutations also adversely affect infectious virus assembly and release, processes in which NS3 also participates.
Methods: We studied the impact of 25 previously identified PI-resistance mutations on the capacity of genotype 1a H77S RNA to replicate in cell culture and produce infectious virus.
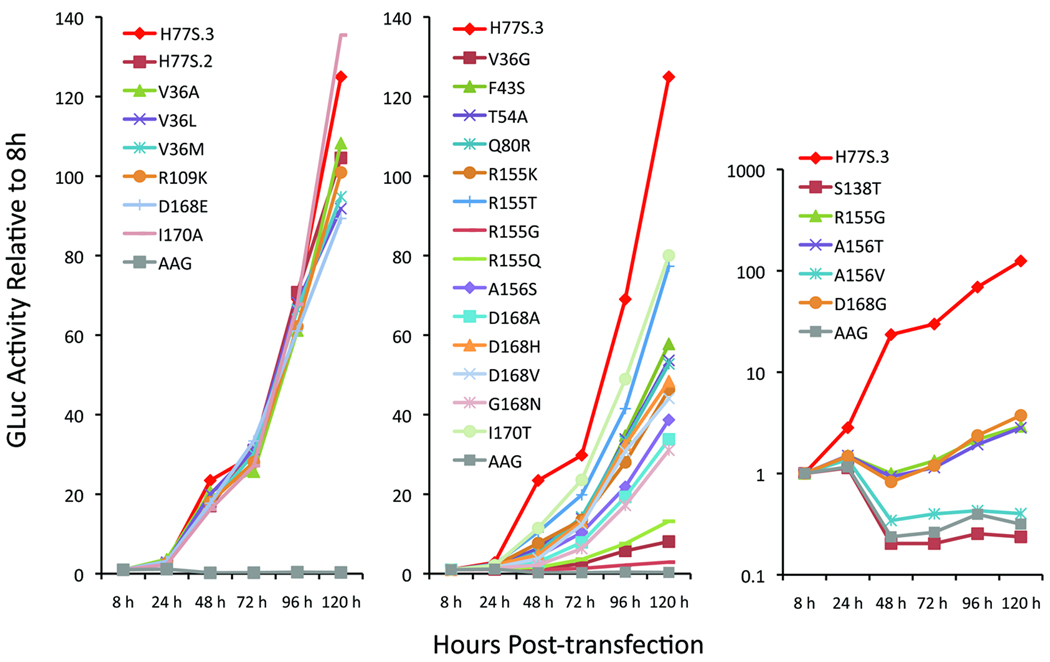
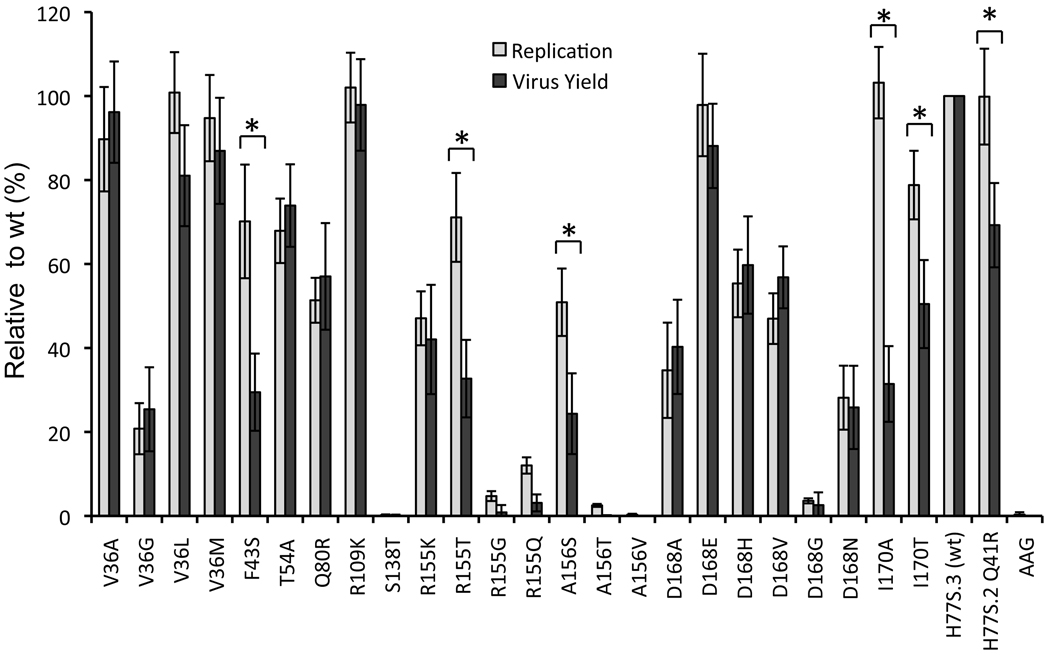
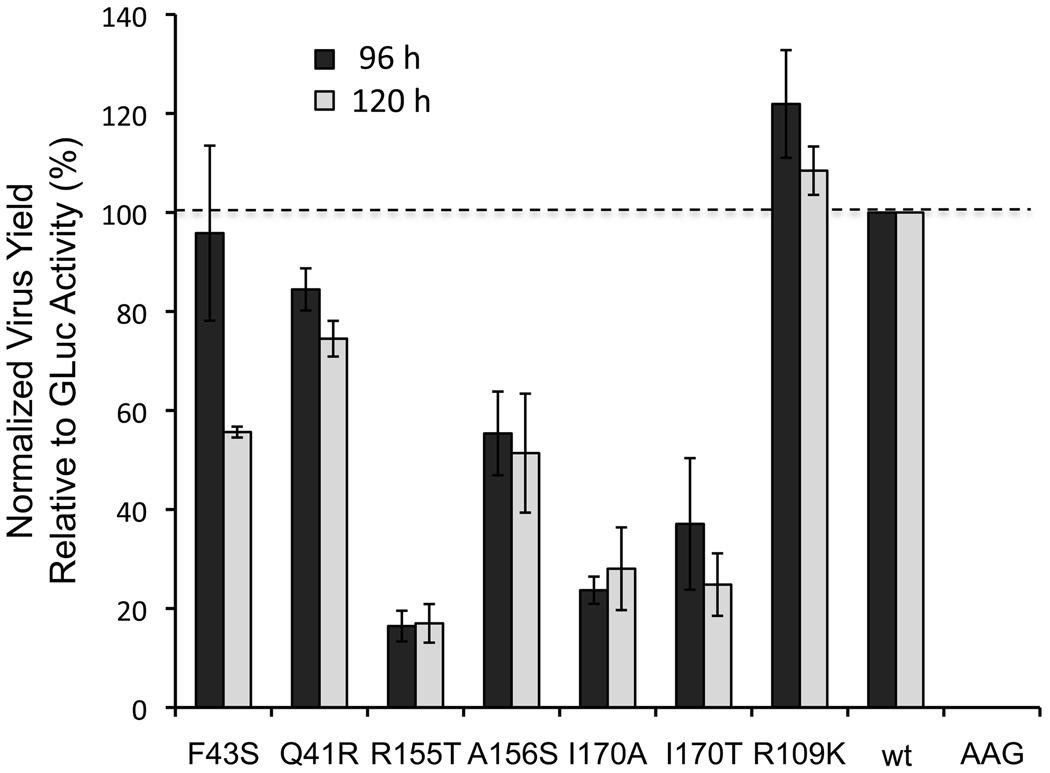
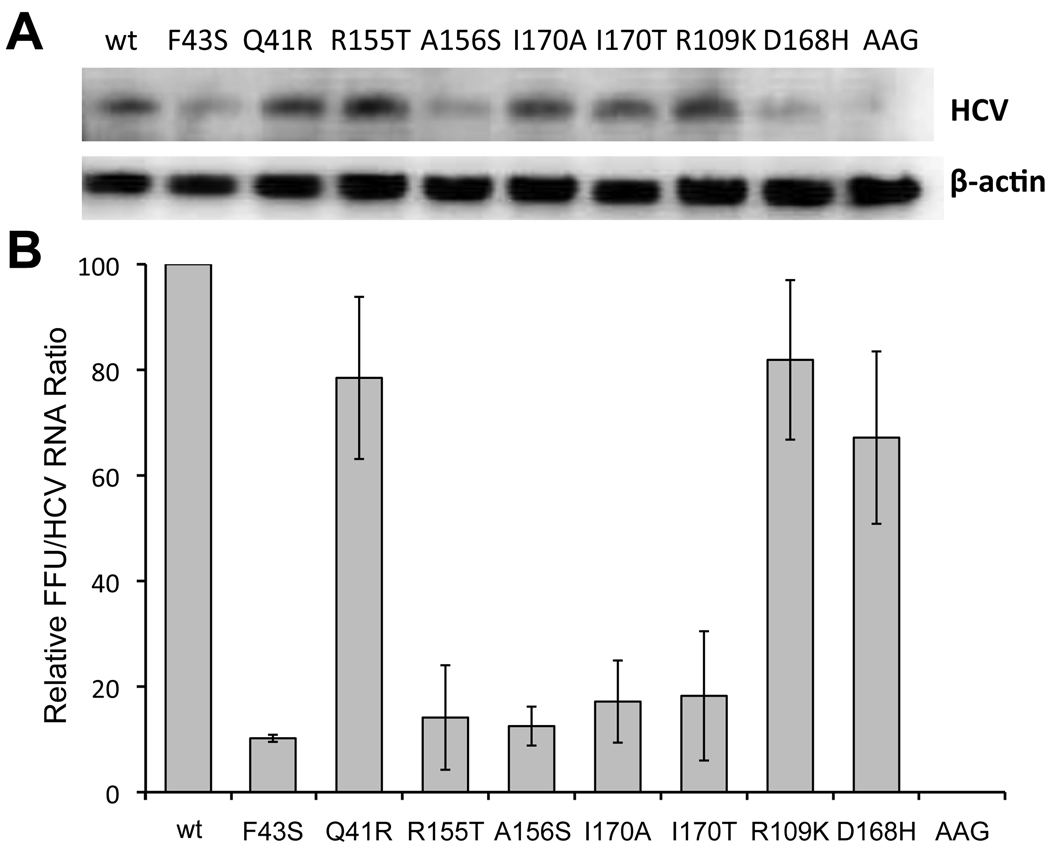
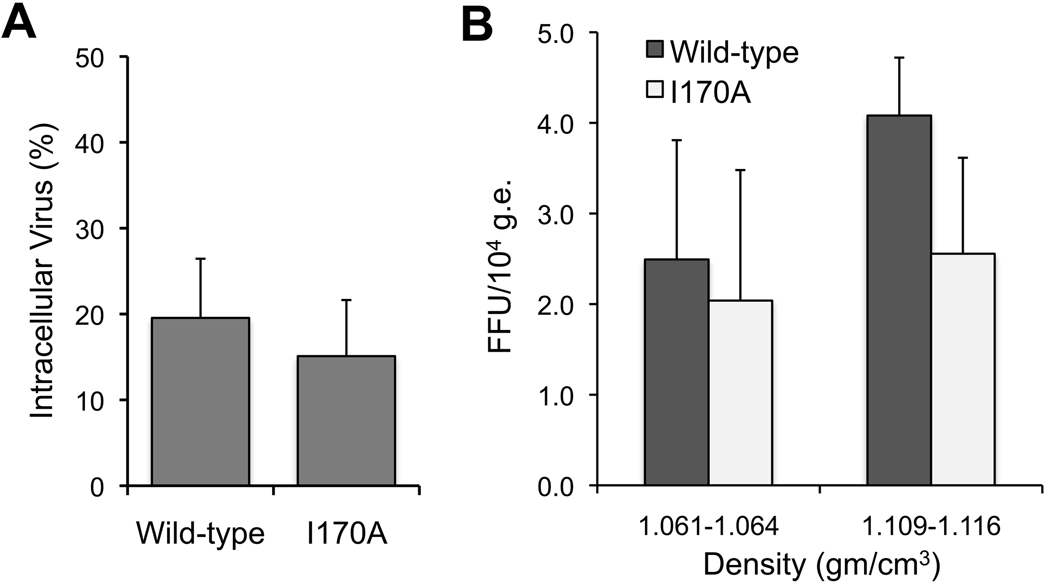
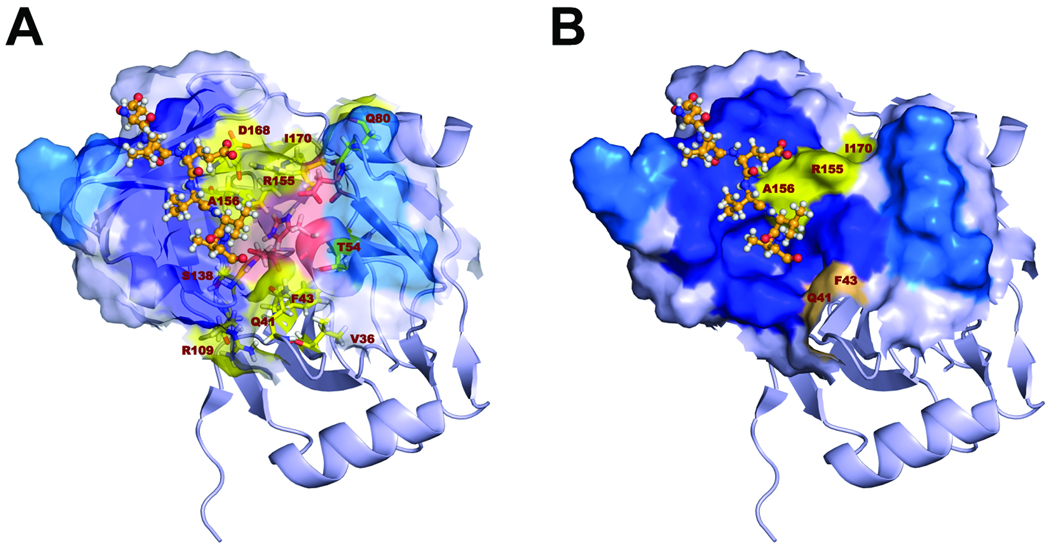
Results: Most PI-resistance mutations resulted in moderate loss of replication competence, although several (V36A/L/M, R109K, and D168E) showed fitness comparable to wild type, whereas others (S138T and A156V) were severely impaired both in RNA replication and infectious virus production. Although reductions in RNA replication capacity correlated with decreased yields of infectious virus for most mutations, a subset of mutants (Q41R, F43S, R155T, A156S, and I170A/T) showed greater impairment in their ability to produce virus than predicted from reductions in RNA replication capacity. Detailed examination of the I170A mutant showed no defect in release of virus from cells and no significant difference in specific infectivity of extracellular virus particles.
Conclusions: Replicon-based assays might underestimate the loss of fitness caused by PI-resistance mutations, because some mutations in the NS3 protease domain specifically impair late steps in the viral life cycle that involve intracellular assembly of infectious virus.
Copyright © 2011 AGA Institute. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Fried M, Shiffman M, Reddy K, et al. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N Engl J Med. 2002;347:975–982. - PubMed
-
- McHutchison J, Everson G, Gordon S, et al. Telaprevir with peginterferon and ribavirin for chronic HCV genotype 1 infection. N Engl J Med. 2009;360:1827–1838. - PubMed
-
- Hézode C, Forestier N, Dusheiko G, et al. Telaprevir and peginterferon with or without ribavirin for chronic HCV infection. N Engl J Med. 2009;360:1839–1850. - PubMed
-
- Sarrazin C, Rouzier R, Wagner F, et al. SCH 503034, a novel hepatitis C virus protease inhibitor, plus pegylated interferon alpha-2b for genotype 1 nonresponders. Gastroenterology. 2007;132:1270–1278. - PubMed
-
- Reesink H, Fanning G, Farha K, et al. Rapid HCV-RNA decline with once daily TMC435: a phase I study in healthy volunteers and hepatitis C patients. Gastroenterology. 2010;138:913–921. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
