

Archaeal-like chaperonins in bacteria
- PMID: 21057109
- PMCID: PMC2996707
- DOI: 10.1073/pnas.1004783107
Archaeal-like chaperonins in bacteria
Abstract
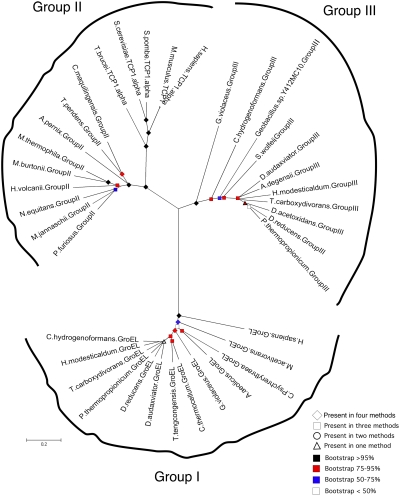
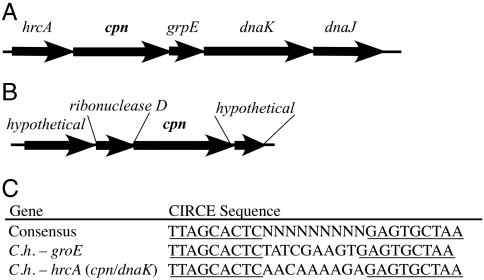
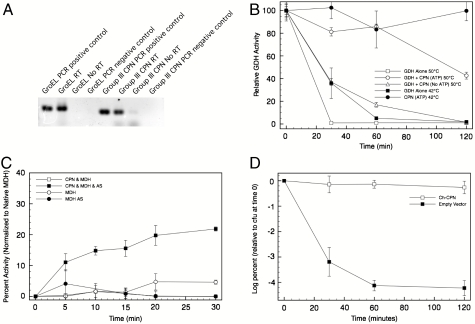
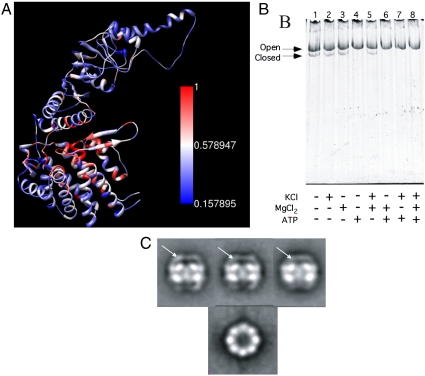
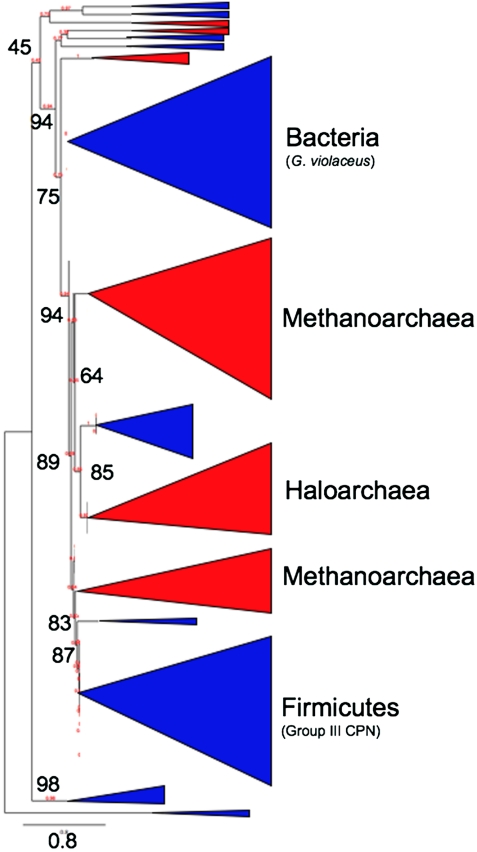
Chaperonins (CPN) are ubiquitous oligomeric protein machines that mediate the ATP-dependent folding of polypeptide chains. These chaperones have not only been assigned stress response and normal housekeeping functions but also have a role in certain human disease states. A longstanding convention divides CPNs into two groups that share many conserved sequence motifs but differ in both structure and distribution. Group I complexes are the well known GroEL/ES heat-shock proteins in bacteria, that also occur in some species of mesophilic archaea and in the endosymbiotic organelles of eukaryotes. Group II CPNs are found only in the cytosol of archaea and eukaryotes. Here we report a third, divergent group of CPNs found in several species of bacteria. We propose to name these Group III CPNs because of their distant relatedness to both Group I and II CPNs as well as their unique genomic context, within the hsp70 operon. The prototype Group III CPN, Carboxydothermus hydrogenoformans chaperonin (Ch-CPN), is able to refold denatured proteins in an ATP-dependent manner and is structurally similar to the Group II CPNs, forming a 16-mer with each subunit contributing to a flexible lid domain. The Group III CPN represent a divergent group of bacterial CPNs distinct from the GroEL/ES CPN found in all bacteria. The Group III lineage may represent an ancient horizontal gene transfer from an archaeon into an early Firmicute lineage. An analysis of their functional and structural characteristics may provide important insights into the early history of this ubiquitous family of proteins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Laksanalamai P, Robb FT. Small heat shock proteins from extremophiles: a review. Extremophiles. 2004;8:1–11. - PubMed
-
- Laksanalamai P, Whitehead TA, Robb FT. Minimal protein-folding systems in hyperthermophilic archaea. Nat Rev Microbiol. 2004;2:315–324. - PubMed
-
- Large AT, Goldberg MD, Lund PA. Chaperones and protein folding in the archaea. Biochem Soc Trans. 2009;37:46–51. - PubMed
-
- Large AT, Lund PA. Archaeal chaperonins. Front Biosci. 2009;14:1304–1324. - PubMed
-
- Macario AJ, Conway De Macario E. The molecular chaperone system and other anti-stress mechanisms in archaea. Front Biosci. 2001;6:D262–283. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
