

The structure of an Iws1/Spt6 complex reveals an interaction domain conserved in TFIIS, Elongin A and Med26
- PMID: 21057455
- PMCID: PMC3020637
- DOI: 10.1038/emboj.2010.272
The structure of an Iws1/Spt6 complex reveals an interaction domain conserved in TFIIS, Elongin A and Med26
Abstract
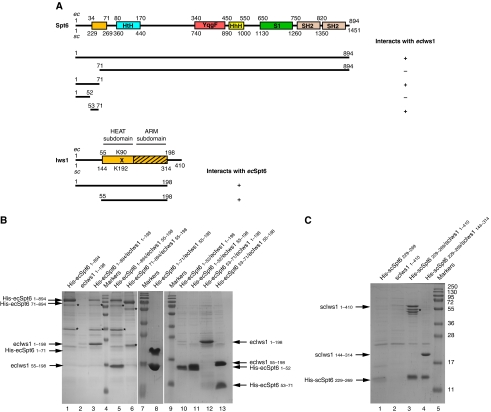
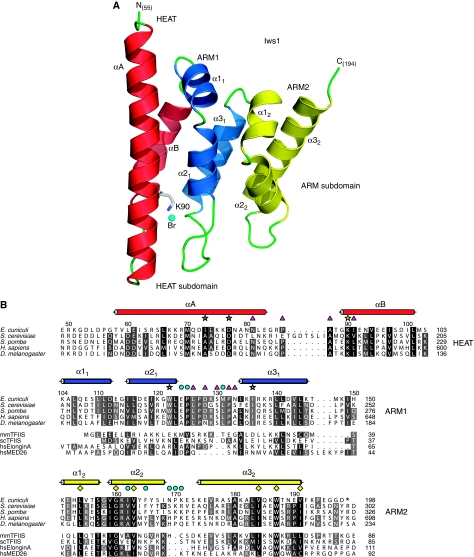
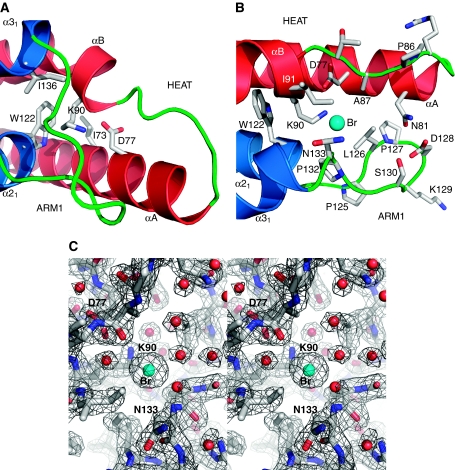
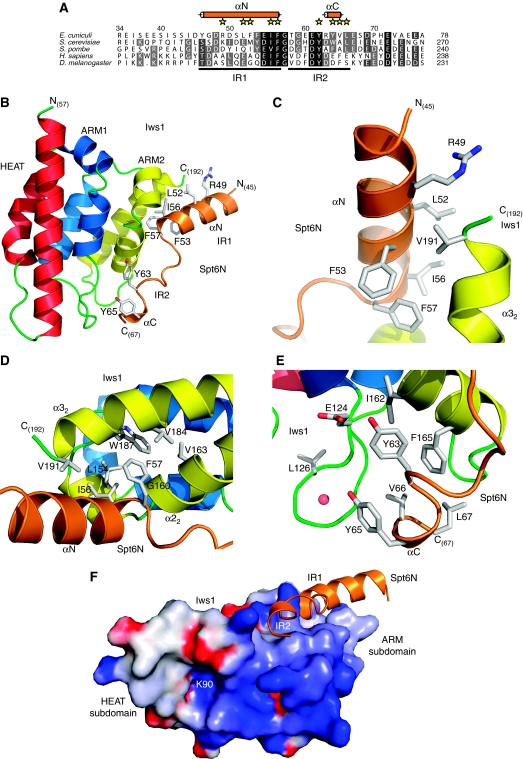
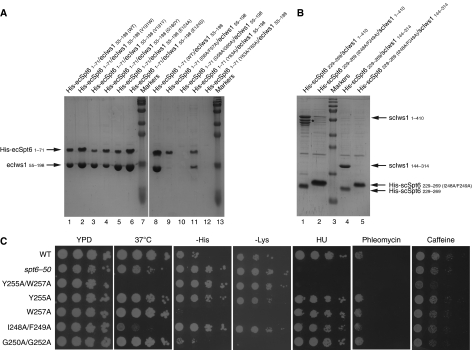
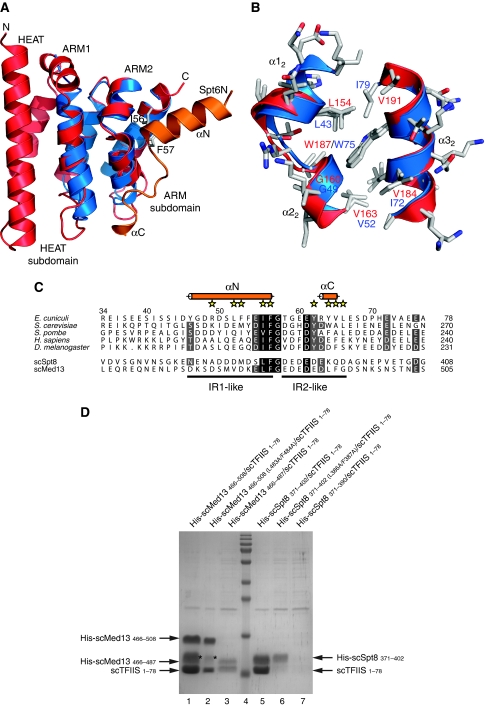
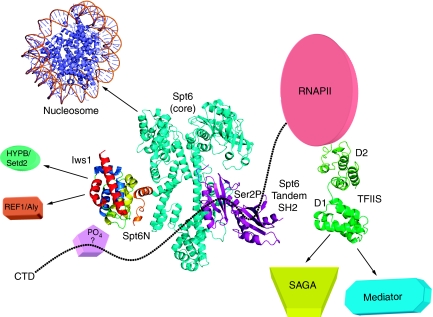
Binding of elongation factor Spt6 to Iws1 provides an effective means for coupling eukaryotic mRNA synthesis, chromatin remodelling and mRNA export. We show that an N-terminal region of Spt6 (Spt6N) is responsible for interaction with Iws1. The crystallographic structures of Encephalitozoon cuniculi Iws1 and the Iws1/Spt6N complex reveal two conserved binding subdomains in Iws1. The first subdomain (one HEAT repeat; HEAT subdomain) is a putative phosphoprotein-binding site most likely involved in an Spt6-independent function of Iws1. The second subdomain (two ARM repeats; ARM subdomain) specifically recognizes a bipartite N-terminal region of Spt6. Mutations that alter this region of Spt6 cause severe phenotypes in vivo. Importantly, the ARM subdomain of Iws1 is conserved in several transcription factors, including TFIIS, Elongin A and Med26. We show that the homologous region in yeast TFIIS enables this factor to interact with SAGA and the Mediator subunits Spt8 and Med13, suggesting the molecular basis for TFIIS recruitment at promoters. Taken together, our results provide new structural information about the Iws1/Spt6 complex and reveal a novel interaction domain used for the formation of transcription networks.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Crystallization and preliminary crystallographic analysis of eukaryotic transcription and mRNA export factor Iws1 from Encephalitozoon cuniculi.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2010 Feb 1;66(Pt 2):207-10. doi: 10.1107/S1744309109052749. Epub 2010 Jan 28. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2010. PMID: 20124725 Free PMC article.
-
Structure of a conserved domain common to the transcription factors TFIIS, elongin A, and CRSP70.J Biol Chem. 2000 Oct 6;275(40):31266-8. doi: 10.1074/jbc.M002595200. J Biol Chem. 2000. PMID: 10811649
-
The TFIIS N-terminal domain (TND): a transcription assembly module at the interface of order and disorder.Biochem Soc Trans. 2023 Feb 27;51(1):125-135. doi: 10.1042/BST20220342. Biochem Soc Trans. 2023. PMID: 36651856 Free PMC article. Review.
-
A ubiquitous disordered protein interaction module orchestrates transcription elongation.Science. 2021 Nov 26;374(6571):1113-1121. doi: 10.1126/science.abe2913. Epub 2021 Nov 25. Science. 2021. PMID: 34822292 Free PMC article.
-
RNA polymerase II elongation factors of Saccharomyces cerevisiae: a targeted proteomics approach.Mol Cell Biol. 2002 Oct;22(20):6979-92. doi: 10.1128/MCB.22.20.6979-6992.2002. Mol Cell Biol. 2002. PMID: 12242279 Free PMC article.
Cited by
-
A conserved genetic interaction between Spt6 and Set2 regulates H3K36 methylation.Nucleic Acids Res. 2019 May 7;47(8):3888-3903. doi: 10.1093/nar/gkz119. Nucleic Acids Res. 2019. PMID: 30793188 Free PMC article.
-
High nitrogen insensitive 9 (HNI9)-mediated systemic repression of root NO3- uptake is associated with changes in histone methylation.Proc Natl Acad Sci U S A. 2011 Aug 9;108(32):13329-34. doi: 10.1073/pnas.1017863108. Epub 2011 Jul 25. Proc Natl Acad Sci U S A. 2011. PMID: 21788519 Free PMC article.
-
Spt6 Is Required for the Fidelity of Promoter Selection.Mol Cell. 2018 Nov 15;72(4):687-699.e6. doi: 10.1016/j.molcel.2018.09.005. Epub 2018 Oct 11. Mol Cell. 2018. PMID: 30318445 Free PMC article.
-
Spt6 regulates intragenic and antisense transcription, nucleosome positioning, and histone modifications genome-wide in fission yeast.Mol Cell Biol. 2013 Dec;33(24):4779-92. doi: 10.1128/MCB.01068-13. Epub 2013 Oct 7. Mol Cell Biol. 2013. PMID: 24100010 Free PMC article.
-
Molecular docking and biological evaluation of a novel IWS1 inhibitor for the treatment of human retroperitoneal liposarcoma.Sci Rep. 2025 Jul 2;15(1):22965. doi: 10.1038/s41598-025-07215-y. Sci Rep. 2025. PMID: 40594510 Free PMC article.
References
-
- Adkins MW, Tyler JK (2006) Transcriptional activators are dispensable for transcription in the absence of Spt6-mediated chromatin reassembly of promoter regions. Mol Cell 21: 405–416 - PubMed
-
- Andrade MA, Petosa C, O'Donoghue SI, Muller CW, Bork P (2001) Comparison of ARM and HEAT protein repeats. J Mol Biol 309: 1–18 - PubMed
-
- Boeke JD, LaCroute F, Fink GR (1984) A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol Gen Genet 197: 345–346 - PubMed
-
- Bond CS, Schuttelkopf AW (2009) ALINE: a WYSIWYG protein-sequence alignment editor for publication-quality alignments. Acta Crystallogr D Biol Crystallogr 65(Part 5): 510–512 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
