

Structural origins of high apparent dielectric constants experienced by ionizable groups in the hydrophobic core of a protein
- PMID: 21059359
- PMCID: PMC3373013
- DOI: 10.1016/j.jmb.2010.10.001
Structural origins of high apparent dielectric constants experienced by ionizable groups in the hydrophobic core of a protein
Abstract
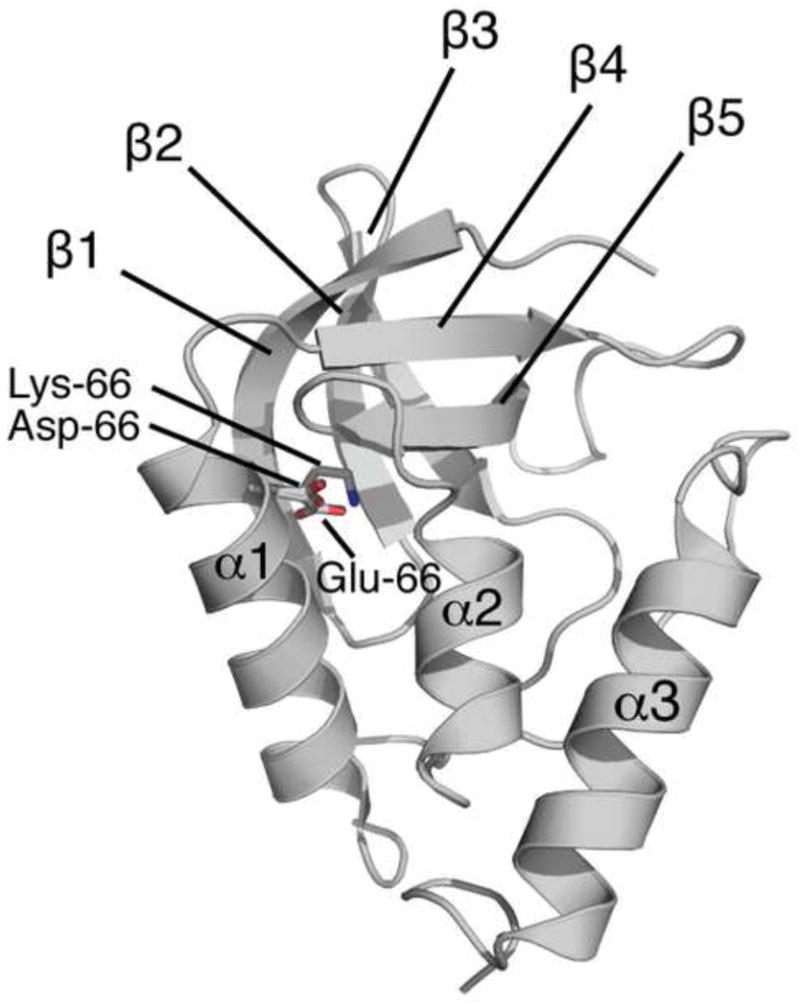
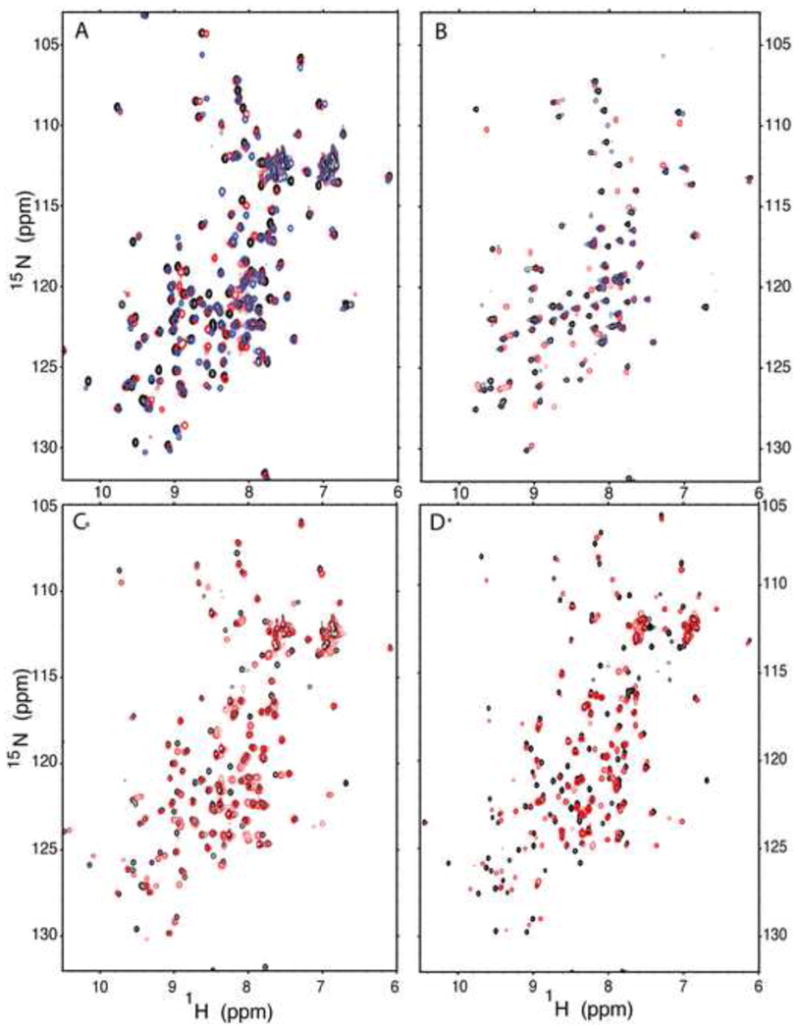
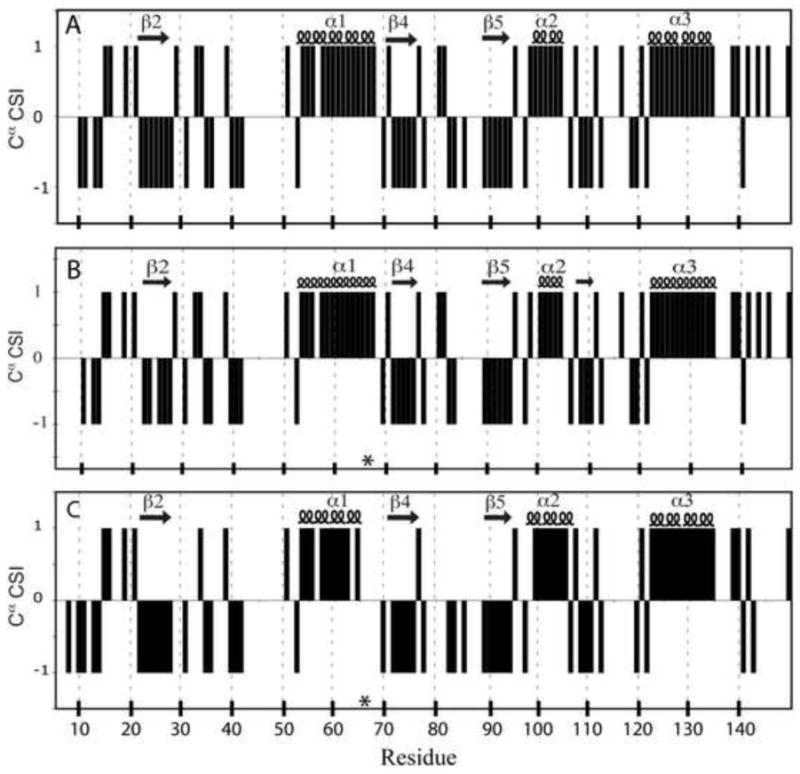
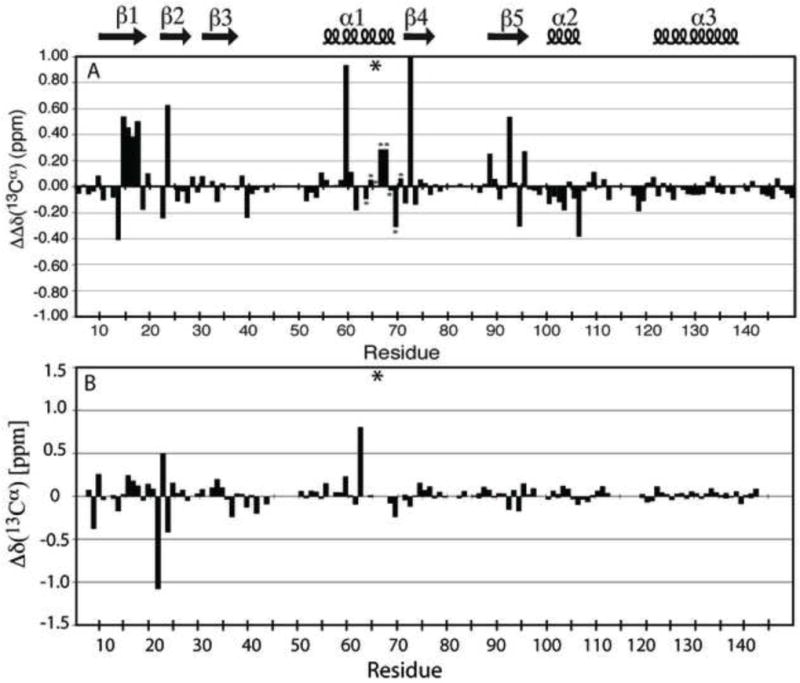
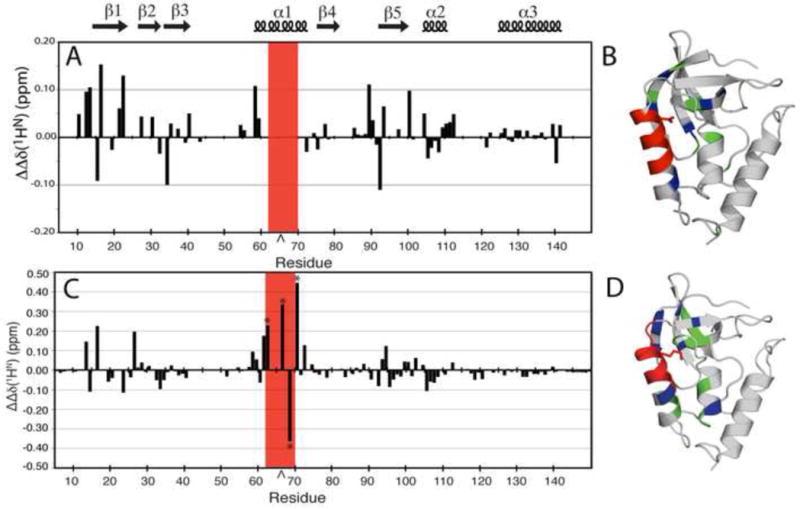
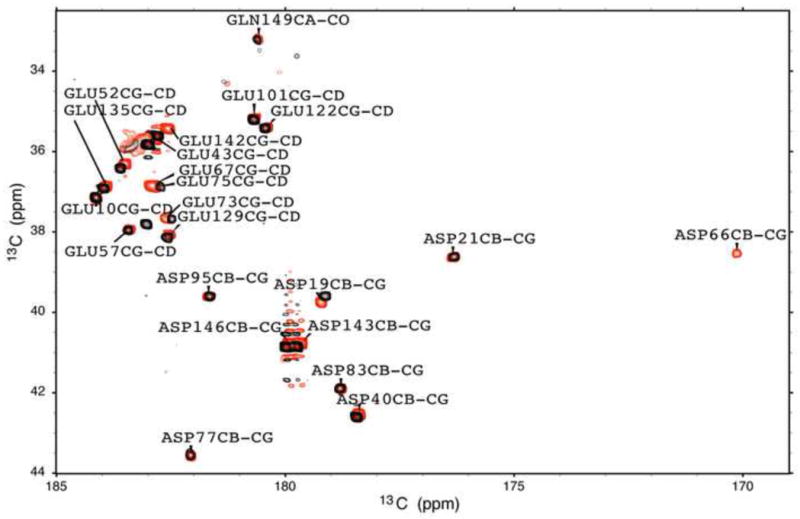
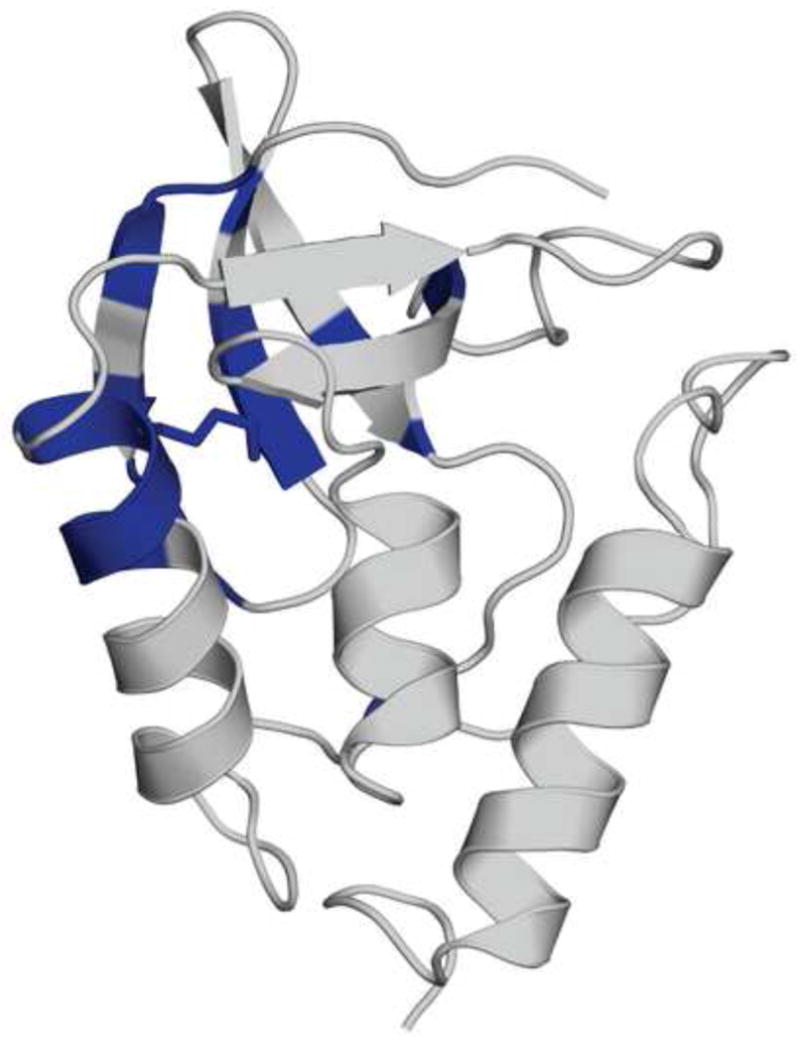
The side chains of Lys66, Asp66, and Glu66 in staphylococcal nuclease are fully buried and surrounded mainly by hydrophobic matter, except for internal water molecules associated with carboxylic oxygen atoms. These ionizable side chains titrate with pK(a) values of 5.7, 8.8, and 8.9, respectively. To reproduce these pK(a) values with continuum electrostatics calculations, we treated the protein with high dielectric constants. We have examined the structural origins of these high apparent dielectric constants by using NMR spectroscopy to characterize the structural response to the ionization of these internal side chains. Substitution of Val66 with Lys66 and Asp66 led to increased conformational fluctuations of the microenvironments surrounding these groups, even under pH conditions where Lys66 and Asp66 are neutral. When Lys66, Asp66, and Glu66 are charged, the proteins remain almost fully folded, but resonances for a few backbone amides adjacent to the internal ionizable residues are broadened. This suggests that the ionization of the internal groups promotes a local increase in dynamics on the intermediate timescale, consistent with either partial unfolding or increased backbone fluctuations of helix 1 near residue 66, or, less likely, with increased fluctuations of the charged side chains at position 66. These experiments confirm that the high apparent dielectric constants reported by internal Lys66, Asp66, and Glu66 reflect localized changes in conformational fluctuations without incurring detectable global structural reorganization. To improve structure-based pK(a) calculations in proteins, we will need to learn how to treat this coupling between ionization of internal groups and local changes in conformational fluctuations explicitly.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Lanyi JK, Luecke H. Bacteriorhodopsin. Current Opinion in Structural Biology. 2001;11:415–419. - PubMed
-
- Rastogi VK, Girvin ME. Structural changes linked to proton translocation by subunit c of the ATP synthase. Nature. 1999;402:263–268. - PubMed
-
- Xie AH, Kelemen L, Hendriks J, White BJ, Hellingwerf KJ, Hoff WD. Formation of a new buried charge drives a large-amplitude protein quake in photoreceptor activation. Biochemistry. 2001;40:1510–1517. - PubMed
-
- Deisenhofer J, Epp O, Miki K, Huber R, Michel H. Structure of the protein subunits in the photosynthetic reaction centre of Rhodopseudomonas viridis at 3[angst] resolution. Nature. 1985;318:618–624. - PubMed
-
- Abramson J, Smirnova I, Kasho V, Verner G, Kaback HR, Iwata S. Structure and Mechanism of the Lactose Permease of Escherichia coli. Science. 2003;301:610. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
