

Different Mi-2 complexes for various developmental functions in Caenorhabditis elegans
- PMID: 21060680
- PMCID: PMC2965115
- DOI: 10.1371/journal.pone.0013681
Different Mi-2 complexes for various developmental functions in Caenorhabditis elegans
Abstract
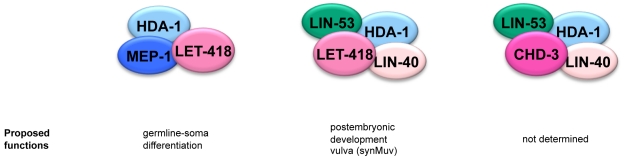
Biochemical purifications from mammalian cells and Xenopus oocytes revealed that vertebrate Mi-2 proteins reside in multisubunit NuRD (Nucleosome Remodeling and Deacetylase) complexes. Since all NuRD subunits are highly conserved in the genomes of C. elegans and Drosophila, it was suggested that NuRD complexes also exist in invertebrates. Recently, a novel dMec complex, composed of dMi-2 and dMEP-1 was identified in Drosophila. The genome of C. elegans encodes two highly homologous Mi-2 orthologues, LET-418 and CHD-3. Here we demonstrate that these proteins define at least three different protein complexes, two distinct NuRD complexes and one MEC complex. The two canonical NuRD complexes share the same core subunits HDA-1/HDAC, LIN-53/RbAp and LIN-40/MTA, but differ in their Mi-2 orthologues LET-418 or CHD-3. LET-418 but not CHD-3, interacts with the Krüppel-like protein MEP-1 in a distinct complex, the MEC complex. Based on microarrays analyses, we propose that MEC constitutes an important LET-418 containing regulatory complex during C. elegans embryonic and early larval development. It is required for the repression of germline potential in somatic cells and acts when blastomeres are still dividing and differentiating. The two NuRD complexes may not be important for the early development, but may act later during postembryonic development. Altogether, our data suggest a considerable complexity in the composition, the developmental function and the tissue-specificity of the different C. elegans Mi-2 complexes.
Conflict of interest statement
Figures



Similar articles
-
Novel Mi-2 related ATP-dependent chromatin remodelers.Epigenetics. 2009 May 16;4(4):209-11. doi: 10.4161/epi.8933. Epub 2009 May 6. Epigenetics. 2009. PMID: 19535903 Review.
-
dMec: a novel Mi-2 chromatin remodelling complex involved in transcriptional repression.EMBO J. 2009 Mar 4;28(5):533-44. doi: 10.1038/emboj.2009.3. Epub 2009 Jan 22. EMBO J. 2009. PMID: 19165147 Free PMC article.
-
MEP-1 and a homolog of the NURD complex component Mi-2 act together to maintain germline-soma distinctions in C. elegans.Cell. 2002 Dec 27;111(7):991-1002. doi: 10.1016/s0092-8674(02)01202-3. Cell. 2002. PMID: 12507426
-
A Network of Chromatin Factors Is Regulating the Transition to Postembryonic Development in Caenorhabditis elegans.G3 (Bethesda). 2017 Feb 9;7(2):343-353. doi: 10.1534/g3.116.037747. G3 (Bethesda). 2017. PMID: 28007841 Free PMC article.
-
Mi-2/NuRD: multiple complexes for many purposes.Biochim Biophys Acta. 2004 Mar 15;1677(1-3):52-7. doi: 10.1016/j.bbaexp.2003.10.010. Biochim Biophys Acta. 2004. PMID: 15020045 Review.
Cited by
-
NuRD chromatin remodeling is required to repair exogenous DSBs in the Caenorhabditis elegans germline.bioRxiv [Preprint]. 2024 Sep 15:2024.09.14.613027. doi: 10.1101/2024.09.14.613027. bioRxiv. 2024. PMID: 39314477 Free PMC article. Preprint.
-
Affinity-based enrichment strategies to assay methyl-CpG binding activity and DNA methylation in early Xenopus embryos.BMC Res Notes. 2011 Aug 18;4:300. doi: 10.1186/1756-0500-4-300. BMC Res Notes. 2011. PMID: 21851637 Free PMC article.
-
A team of heterochromatin factors collaborates with small RNA pathways to combat repetitive elements and germline stress.Elife. 2017 Mar 15;6:e21666. doi: 10.7554/eLife.21666. Elife. 2017. PMID: 28294943 Free PMC article.
-
BLIMP-1/BLMP-1 and Metastasis-Associated Protein Regulate Stress Resistant Development in Caenorhabditis elegans.Genetics. 2016 Aug;203(4):1721-32. doi: 10.1534/genetics.116.190793. Epub 2016 Jun 22. Genetics. 2016. PMID: 27334271 Free PMC article.
-
CHD chromatin remodelers and the transcription cycle.Transcription. 2011 Nov-Dec;2(6):244-53. doi: 10.4161/trns.2.6.17840. Epub 2011 Nov 1. Transcription. 2011. PMID: 22223048 Free PMC article. Review.
References
-
- Seelig HP, Moosbrugger I, Ehrfeld H, Fink T, Renz M, et al. The major dermatomyositis-specific Mi-2 autoantigen is a presumed helicase involved in transcriptional activation. Arthritis Rheum. 1995;38:1389–1399. - PubMed
-
- Ahringer J. NuRD and SIN3 histone deacetylase complexes in development. Trends Genet. 2000;16:351–356. - PubMed
-
- Bowen NJ, Fujita N, Kajita M, Wade PA. Mi-2/NuRD: multiple complexes for many purposes. Biochim Biophys Acta. 2004;1677:52–57. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
