

Facioscapulohumeral dystrophy: incomplete suppression of a retrotransposed gene
- PMID: 21060811
- PMCID: PMC2965761
- DOI: 10.1371/journal.pgen.1001181
Facioscapulohumeral dystrophy: incomplete suppression of a retrotransposed gene
Abstract
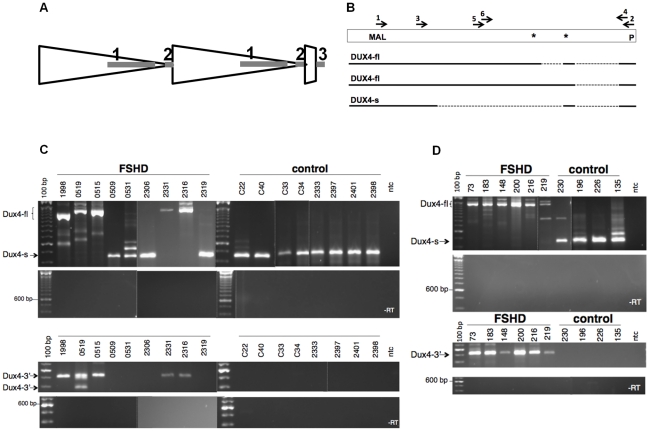
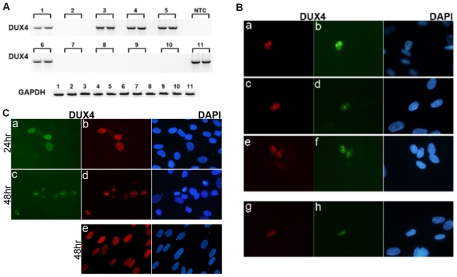
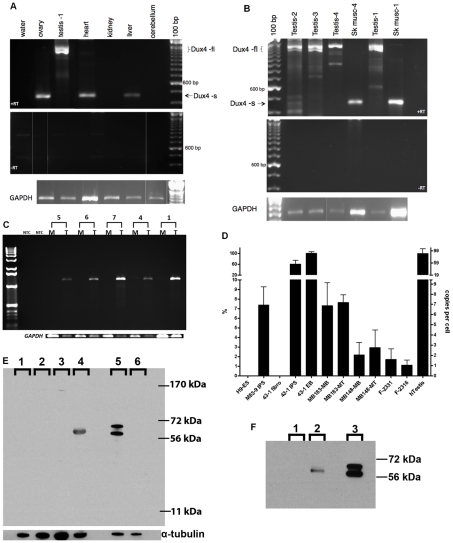
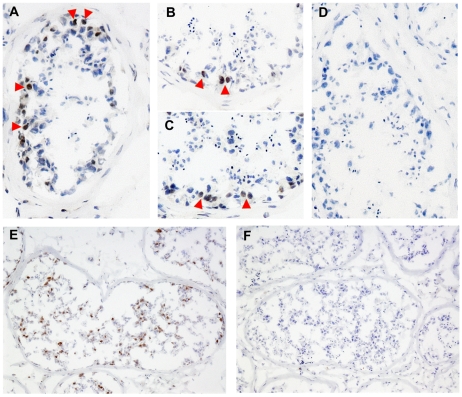
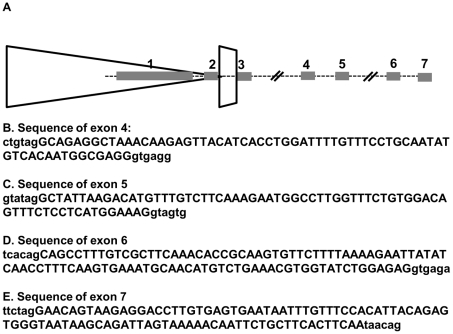
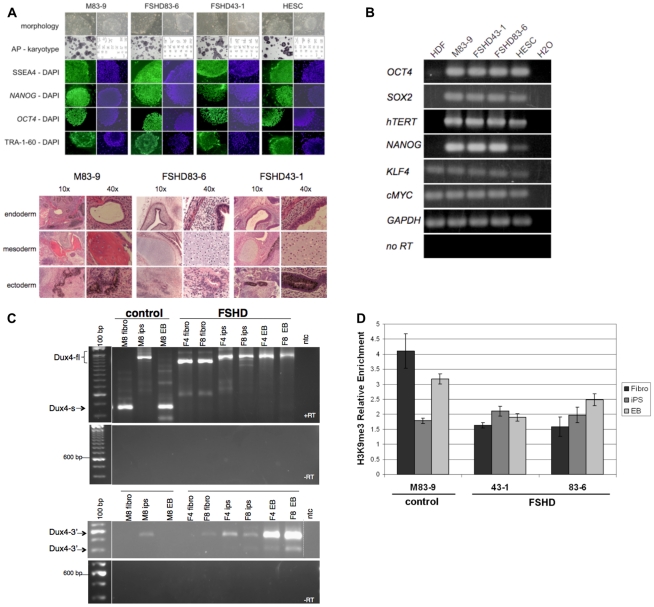
Each unit of the D4Z4 macrosatellite repeat contains a retrotransposed gene encoding the DUX4 double-homeobox transcription factor. Facioscapulohumeral dystrophy (FSHD) is caused by deletion of a subset of the D4Z4 units in the subtelomeric region of chromosome 4. Although it has been reported that the deletion of D4Z4 units induces the pathological expression of DUX4 mRNA, the association of DUX4 mRNA expression with FSHD has not been rigorously investigated, nor has any human tissue been identified that normally expresses DUX4 mRNA or protein. We show that FSHD muscle expresses a different splice form of DUX4 mRNA compared to control muscle. Control muscle produces low amounts of a splice form of DUX4 encoding only the amino-terminal portion of DUX4. FSHD muscle produces low amounts of a DUX4 mRNA that encodes the full-length DUX4 protein. The low abundance of full-length DUX4 mRNA in FSHD muscle cells represents a small subset of nuclei producing a relatively high abundance of DUX4 mRNA and protein. In contrast to control skeletal muscle and most other somatic tissues, full-length DUX4 transcript and protein is expressed at relatively abundant levels in human testis, most likely in the germ-line cells. Induced pluripotent (iPS) cells also express full-length DUX4 and differentiation of control iPS cells to embryoid bodies suppresses expression of full-length DUX4, whereas expression of full-length DUX4 persists in differentiated FSHD iPS cells. Together, these findings indicate that full-length DUX4 is normally expressed at specific developmental stages and is suppressed in most somatic tissues. The contraction of the D4Z4 repeat in FSHD results in a less efficient suppression of the full-length DUX4 mRNA in skeletal muscle cells. Therefore, FSHD represents the first human disease to be associated with the incomplete developmental silencing of a retrogene array normally expressed early in development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
FSHD: a repeat contraction disease finally ready to expand (our understanding of its pathogenesis).PLoS Genet. 2010 Oct 28;6(10):e1001180. doi: 10.1371/journal.pgen.1001180. PLoS Genet. 2010. PMID: 21060814 Free PMC article. No abstract available.
References
-
- Tawil R, Van Der Maarel SM. Facioscapulohumeral muscular dystrophy. Muscle Nerve. 2006;34:1–15. - PubMed
-
- Zeng W, de Greef JC, Chen YY, Chien R, Kong X, et al. Specific loss of histone H3 lysine 9 trimethylation and HP1gamma/cohesin binding at D4Z4 repeats is associated with facioscapulohumeral dystrophy (FSHD). PLoS Genet. 2009;5:e1000559. doi: 10.1371/journal.pgen.1000559. - DOI - PMC - PubMed
-
- van Overveld PG, Lemmers RJ, Sandkuijl LA, Enthoven L, Winokur ST, et al. Hypomethylation of D4Z4 in 4q-linked and non-4q-linked facioscapulohumeral muscular dystrophy. Nat Genet. 2003;35:315–317. - PubMed
-
- de Greef JC, Lemmers RJ, van Engelen BG, Sacconi S, Venance SL, et al. Common epigenetic changes of D4Z4 in contraction-dependent and contraction-independent FSHD. Hum Mutat. 2009;30:1449–1459. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
