

Nuclear export and import of human hepatitis B virus capsid protein and particles
- PMID: 21060813
- PMCID: PMC2965763
- DOI: 10.1371/journal.ppat.1001162
Nuclear export and import of human hepatitis B virus capsid protein and particles
Abstract
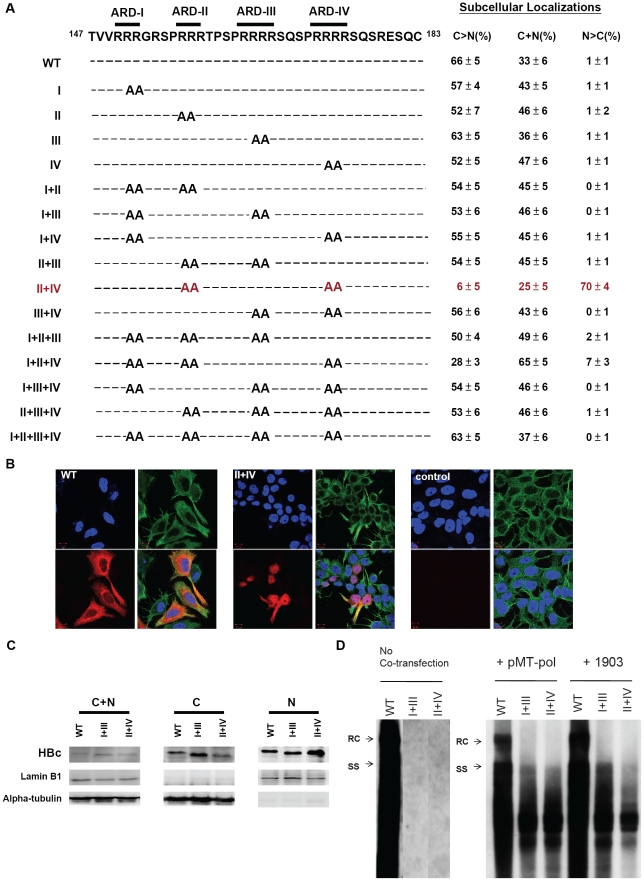
It remains unclear what determines the subcellular localization of hepatitis B virus (HBV) core protein (HBc) and particles. To address this fundamental issue, we have identified four distinct HBc localization signals in the arginine rich domain (ARD) of HBc, using immunofluorescence confocal microscopy and fractionation/Western blot analysis. ARD consists of four tight clustering arginine-rich subdomains. ARD-I and ARD-III are associated with two co-dependent nuclear localization signals (NLS), while ARD-II and ARD-IV behave like two independent nuclear export signals (NES). This conclusion is based on five independent lines of experimental evidence: i) Using an HBV replication system in hepatoma cells, we demonstrated in a double-blind manner that only the HBc of mutant ARD-II+IV, among a total of 15 ARD mutants, can predominantly localize to the nucleus. ii) These results were confirmed using a chimera reporter system by placing mutant or wild type HBc trafficking signals in the heterologous context of SV40 large T antigen (LT). iii) By a heterokaryon or homokaryon analysis, the fusion protein of SV40 LT-HBc ARD appeared to transport from nuclei of transfected donor cells to nuclei of recipient cells, suggesting the existence of an NES in HBc ARD. This putative NES is leptomycin B resistant. iv) We demonstrated by co-immunoprecipitation that HBc ARD can physically interact with a cellular factor TAP/NXF1 (Tip-associated protein/nuclear export factor-1), which is known to be important for nuclear export of mRNA and proteins. Treatment with a TAP-specific siRNA strikingly shifted cytoplasmic HBc to nucleus, and led to a near 7-fold reduction of viral replication, and a near 10-fold reduction in HBsAg secretion. v) HBc of mutant ARD-II+IV was accumulated predominantly in the nucleus in a mouse model by hydrodynamic delivery. In addition to the revised map of NLS, our results suggest that HBc could shuttle rapidly between nucleus and cytoplasm via a novel TAP-dependent NES.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures









Similar articles
-
Nuclear export of human hepatitis B virus core protein and pregenomic RNA depends on the cellular NXF1-p15 machinery.PLoS One. 2014 Oct 31;9(10):e106683. doi: 10.1371/journal.pone.0106683. eCollection 2014. PLoS One. 2014. PMID: 25360769 Free PMC article.
-
A Homokaryon Assay for Nucleocytoplasmic Shuttling Activity of HBV Core Protein.Methods Mol Biol. 2017;1540:53-58. doi: 10.1007/978-1-4939-6700-1_5. Methods Mol Biol. 2017. PMID: 27975307
-
Nucleic acid chaperone activity associated with the arginine-rich domain of human hepatitis B virus core protein.J Virol. 2014 Mar;88(5):2530-43. doi: 10.1128/JVI.03235-13. Epub 2013 Dec 18. J Virol. 2014. PMID: 24352445 Free PMC article.
-
Virion Secretion of Hepatitis B Virus Naturally Occurring Core Antigen Variants.Cells. 2020 Dec 30;10(1):43. doi: 10.3390/cells10010043. Cells. 2020. PMID: 33396864 Free PMC article. Review.
-
The diverse functions of the hepatitis B core/capsid protein (HBc) in the viral life cycle: Implications for the development of HBc-targeting antivirals.Antiviral Res. 2018 Jan;149:211-220. doi: 10.1016/j.antiviral.2017.11.015. Epub 2017 Nov 26. Antiviral Res. 2018. PMID: 29183719 Free PMC article. Review.
Cited by
-
ViralZone: recent updates to the virus knowledge resource.Nucleic Acids Res. 2013 Jan;41(Database issue):D579-83. doi: 10.1093/nar/gks1220. Epub 2012 Nov 28. Nucleic Acids Res. 2013. PMID: 23193299 Free PMC article.
-
Regulation of multiple stages of hepadnavirus replication by the carboxyl-terminal domain of viral core protein in trans.J Virol. 2015 Mar;89(5):2918-30. doi: 10.1128/JVI.03116-14. Epub 2014 Dec 24. J Virol. 2015. PMID: 25540387 Free PMC article.
-
Characterization of the host factors required for hepadnavirus covalently closed circular (ccc) DNA formation.PLoS One. 2012;7(8):e43270. doi: 10.1371/journal.pone.0043270. Epub 2012 Aug 13. PLoS One. 2012. PMID: 22912842 Free PMC article.
-
Use of a Fluorescent Analogue of a HBV Core Protein-Directed Drug To Interrogate an Antiviral Mechanism.J Am Chem Soc. 2018 Nov 14;140(45):15261-15269. doi: 10.1021/jacs.8b07988. Epub 2018 Oct 30. J Am Chem Soc. 2018. PMID: 30375863 Free PMC article.
-
Biology of the hepatitis B virus (HBV) core and capsid assembly modulators (CAMs) for chronic hepatitis B (CHB) cure.Glob Health Med. 2023 Aug 31;5(4):199-207. doi: 10.35772/ghm.2023.01065. Glob Health Med. 2023. PMID: 37655181 Free PMC article. Review.
References
-
- Ganem D, Prince AM. Hepatitis B virus infection–natural history and clinical consequences. N Engl J Med. 2004;350:1118–1129. - PubMed
-
- Chang MH, Chen CJ, Lai MS, Hsu HM, Wu TC, et al. Universal hepatitis B vaccination in Taiwan and the incidence of hepatocellular carcinoma in children. Taiwan Childhood Hepatoma Study Group. N Engl J Med. 1997;336:1855–1859. - PubMed
-
- Shih C. Functional Significance of Naturally Occurring Hepatitis B Virus Variants. In: Locarnini SLC, editor. Human Virus Guides- Human Hepatitis B Viruses. London, UK: International Medical Press; 2008. pp. 23–41.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
