

Nasty viruses, costly plasmids, population dynamics, and the conditions for establishing and maintaining CRISPR-mediated adaptive immunity in bacteria
- PMID: 21060859
- PMCID: PMC2965746
- DOI: 10.1371/journal.pgen.1001171
Nasty viruses, costly plasmids, population dynamics, and the conditions for establishing and maintaining CRISPR-mediated adaptive immunity in bacteria
Abstract
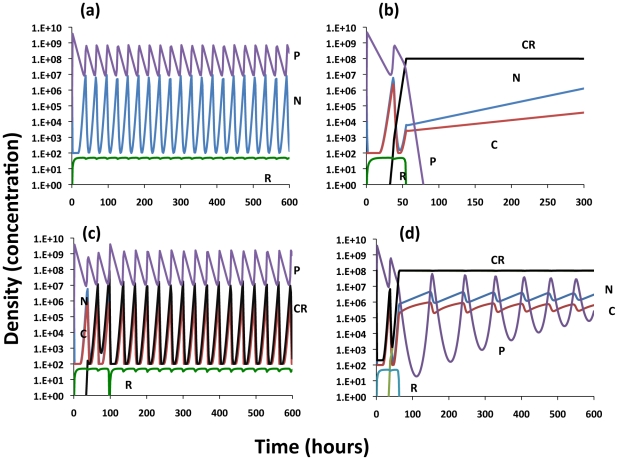
Clustered, Regularly Interspaced Short Palindromic Repeats (CRISPR) abound in the genomes of almost all archaebacteria and nearly half the eubacteria sequenced. Through a genetic interference mechanism, bacteria with CRISPR regions carrying copies of the DNA of previously encountered phage and plasmids abort the replication of phage and plasmids with these sequences. Thus it would seem that protection against infecting phage and plasmids is the selection pressure responsible for establishing and maintaining CRISPR in bacterial populations. But is it? To address this question and provide a framework and hypotheses for the experimental study of the ecology and evolution of CRISPR, I use mathematical models of the population dynamics of CRISPR-encoding bacteria with lytic phage and conjugative plasmids. The results of the numerical (computer simulation) analysis of the properties of these models with parameters in the ranges estimated for Escherichia coli and its phage and conjugative plasmids indicate: (1) In the presence of lytic phage there are broad conditions where bacteria with CRISPR-mediated immunity will have an advantage in competition with non-CRISPR bacteria with otherwise higher Malthusian fitness. (2) These conditions for the existence of CRISPR are narrower when there is envelope resistance to the phage. (3) While there are situations where CRISPR-mediated immunity can provide bacteria an advantage in competition with higher Malthusian fitness bacteria bearing deleterious conjugative plasmids, the conditions for this to obtain are relatively narrow and the intensity of selection favoring CRISPR weak. The parameters of these models can be independently estimated, the assumption behind their construction validated, and the hypotheses generated from the analysis of their properties tested in experimental populations of bacteria with lytic phage and conjugative plasmids. I suggest protocols for estimating these parameters and outline the design of experiments to evaluate the validity of these models and test these hypotheses.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Ochman H, Lawrence JG, Groisman EA. Lateral gene transfer and the nature of bacterial innovation. Nature. 2000;405:299–304. - PubMed
-
- Hall RM. Mobile gene cassettes and integrons: moving antibiotic resistance genes in gram-negative bacteria. Ciba Foundation Symposium. 1997;207:192–202; discussion 202–195. - PubMed
-
- Hall RM. The role of gene cassettes and integrons in the horizontal transfer of genes in Gram-negative bacteria. In: Kado MSaCI., editor. Horizontral Gene Transfer. New York: Chapman & Hall; 1998. pp. 53–62.
-
- Mazel D, Davies J. Gene capture in : Response. Trends Microbiol. 1999;7:95. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
