

Splice isoforms of the polyglutamine disease protein ataxin-3 exhibit similar enzymatic yet different aggregation properties
- PMID: 21060878
- PMCID: PMC2965175
- DOI: 10.1371/journal.pone.0013695
Splice isoforms of the polyglutamine disease protein ataxin-3 exhibit similar enzymatic yet different aggregation properties
Abstract
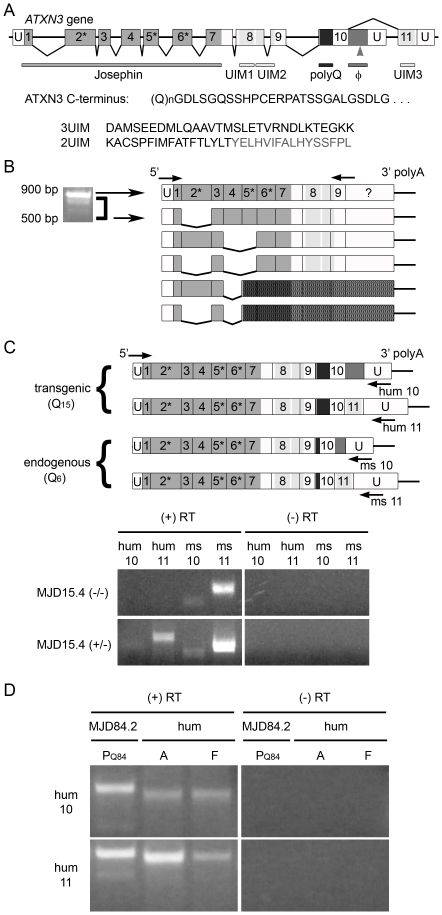
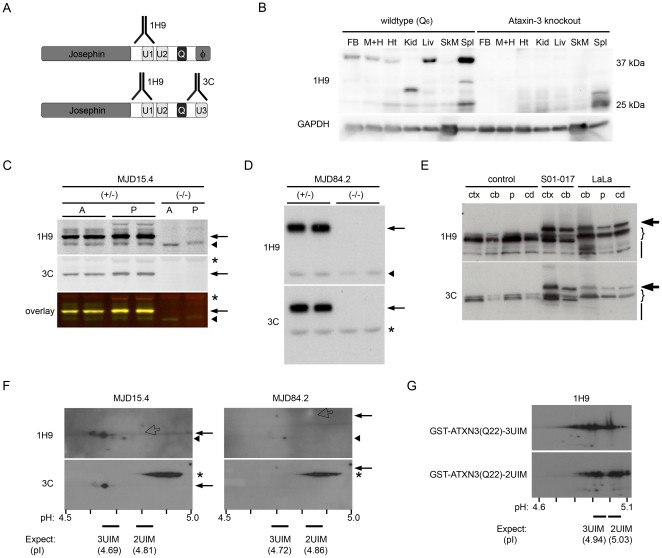
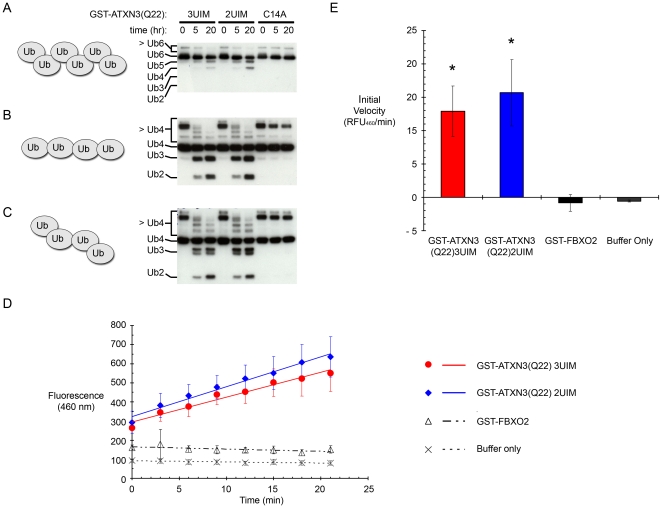
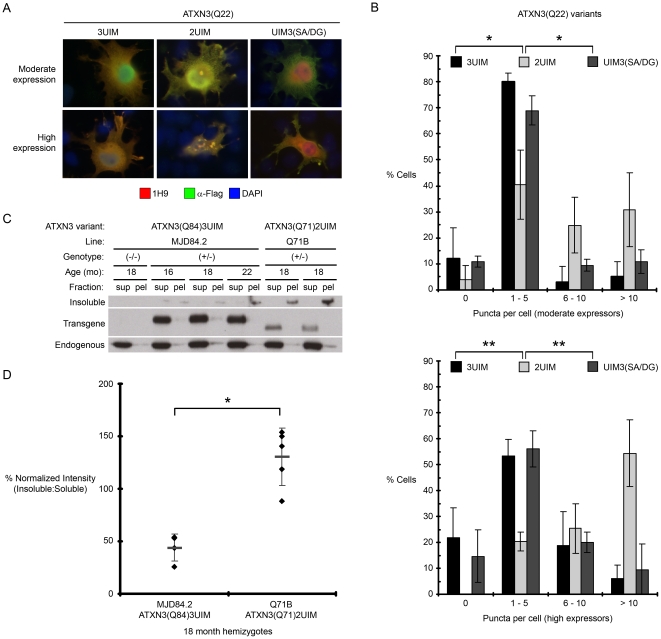
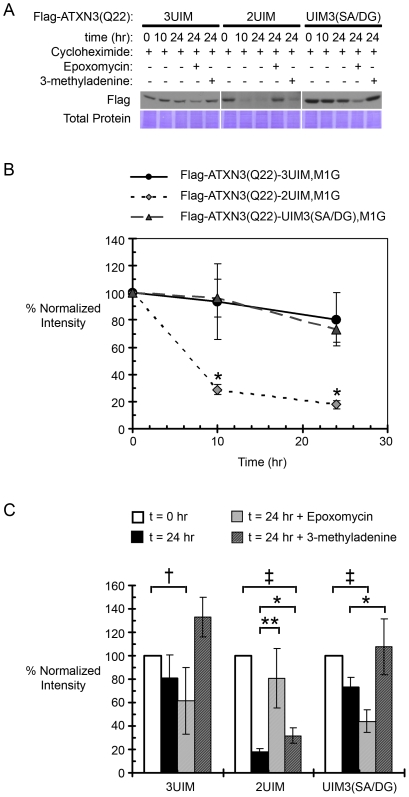
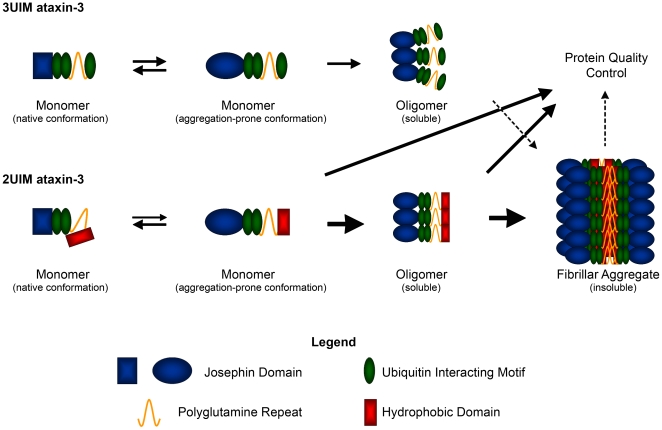
Protein context clearly influences neurotoxicity in polyglutamine diseases, but the contribution of alternative splicing to this phenomenon has rarely been investigated. Ataxin-3, a deubiquitinating enzyme and the disease protein in SCA3, is alternatively spliced to encode either a C-terminal hydrophobic stretch or a third ubiquitin interacting motif (termed 2UIM and 3UIM isoforms, respectively). In light of emerging insights into ataxin-3 function, we examined the significance of this splice variation. We confirmed neural expression of several minor 5' variants and both of the known 3' ataxin-3 splice variants. Regardless of polyglutamine expansion, 3UIM ataxin-3 is the predominant isoform in brain. Although 2UIM and 3UIM ataxin-3 display similar in vitro deubiquitinating activity, 2UIM ataxin-3 is more prone to aggregate and more rapidly degraded by the proteasome. Our data demonstrate how alternative splicing of sequences distinct from the trinucleotide repeat can alter properties of the encoded polyglutamine disease protein and thereby perhaps contribute to selective neurotoxicity.
Conflict of interest statement
Figures
Similar articles
-
A knockin mouse model of spinocerebellar ataxia type 3 exhibits prominent aggregate pathology and aberrant splicing of the disease gene transcript.Hum Mol Genet. 2015 Mar 1;24(5):1211-24. doi: 10.1093/hmg/ddu532. Epub 2014 Oct 15. Hum Mol Genet. 2015. PMID: 25320121 Free PMC article.
-
In vivo suppression of polyglutamine neurotoxicity by C-terminus of Hsp70-interacting protein (CHIP) supports an aggregation model of pathogenesis.Neurobiol Dis. 2009 Mar;33(3):342-53. doi: 10.1016/j.nbd.2008.10.016. Epub 2008 Nov 8. Neurobiol Dis. 2009. PMID: 19084066 Free PMC article.
-
Mouse ataxin-3 functional knock-out model.Neuromolecular Med. 2011 Mar;13(1):54-65. doi: 10.1007/s12017-010-8137-3. Epub 2010 Oct 14. Neuromolecular Med. 2011. PMID: 20945165 Free PMC article.
-
Polyglutamine diseases: the special case of ataxin-3 and Machado-Joseph disease.Prog Neurobiol. 2011 Sep 15;95(1):26-48. doi: 10.1016/j.pneurobio.2011.06.007. Epub 2011 Jun 28. Prog Neurobiol. 2011. PMID: 21740957 Review.
-
Progress in pathogenesis studies of spinocerebellar ataxia type 1.Philos Trans R Soc Lond B Biol Sci. 1999 Jun 29;354(1386):1079-81. doi: 10.1098/rstb.1999.0462. Philos Trans R Soc Lond B Biol Sci. 1999. PMID: 10434309 Free PMC article. Review.
Cited by
-
A knockin mouse model of spinocerebellar ataxia type 3 exhibits prominent aggregate pathology and aberrant splicing of the disease gene transcript.Hum Mol Genet. 2015 Mar 1;24(5):1211-24. doi: 10.1093/hmg/ddu532. Epub 2014 Oct 15. Hum Mol Genet. 2015. PMID: 25320121 Free PMC article.
-
The polyglutamine protein ataxin-3 enables normal growth under heat shock conditions in the methylotrophic yeast Pichia pastoris.Sci Rep. 2017 Oct 17;7(1):13417. doi: 10.1038/s41598-017-13814-1. Sci Rep. 2017. PMID: 29042637 Free PMC article.
-
NeuroGeM, a knowledgebase of genetic modifiers in neurodegenerative diseases.BMC Med Genomics. 2013 Nov 14;6:52. doi: 10.1186/1755-8794-6-52. BMC Med Genomics. 2013. PMID: 24229347 Free PMC article.
-
PolyQ disease: misfiring of a developmental cell death program?Trends Cell Biol. 2013 Apr;23(4):168-74. doi: 10.1016/j.tcb.2012.11.003. Epub 2012 Dec 8. Trends Cell Biol. 2013. PMID: 23228508 Free PMC article. Review.
-
Cell biology of spinocerebellar ataxia.J Cell Biol. 2012 Apr 16;197(2):167-77. doi: 10.1083/jcb.201105092. J Cell Biol. 2012. PMID: 22508507 Free PMC article. Review.
References
-
- Orr HT, Zoghbi HY. Trinucleotide repeat disorders. Annu Rev Neurosci. 2007;30:575–621. - PubMed
-
- Johnson JM, Castle J, Garrett-Engele P, Kan Z, Loerch PM, et al. Genome-wide survey of human alternative pre-mRNA splicing with exon junction microarrays. Science. 2003;302(5653):2141–4. - PubMed
-
- Tsunemi T, Ishikawa K, Jin H, Mizusawa H. Cell-type-specific alternative splicing in spinocerebellar ataxia type 6. Neuroscience Letters. 2008;447(1):78–81. - PubMed
-
- Tadokoro K, Yamazaki-Inoue M, Tachibana M, Fujishiro M, Nagao K, et al. Frequent occurrence of protein isoforms with or without a single amino acid residue by subtle alternative splicing: the case of Gln in DRPLA affects subcellular localization of the products. J Hum Genet. 2005;50(8):382–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
