

Apoptosis regulators Fas and Bim synergistically control T-lymphocyte homeostatic proliferation
- PMID: 21061436
- PMCID: PMC3334341
- DOI: 10.1002/eji.201040577
Apoptosis regulators Fas and Bim synergistically control T-lymphocyte homeostatic proliferation
Abstract
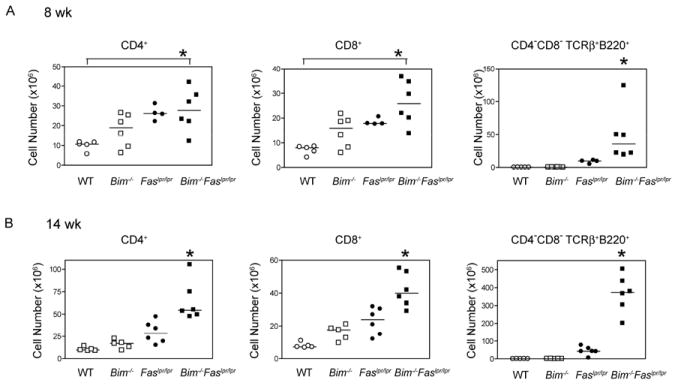
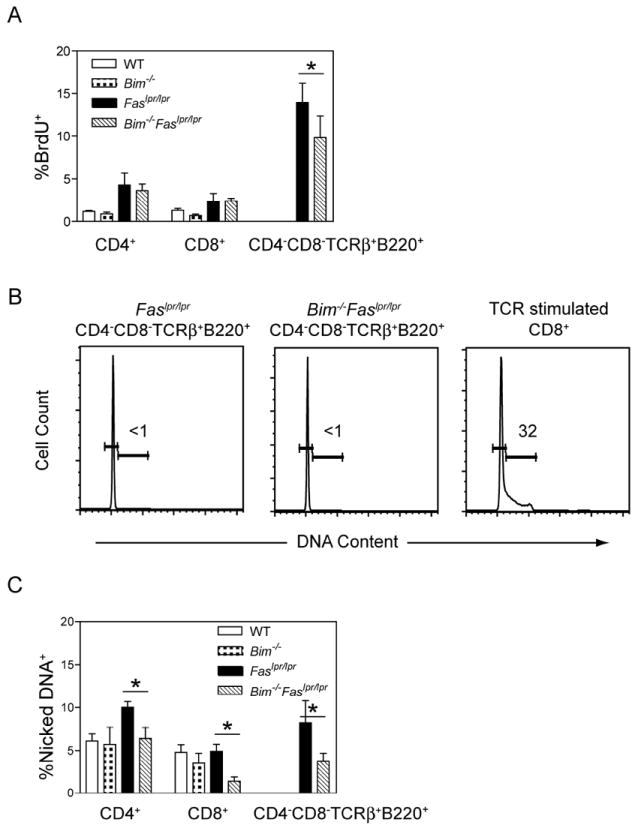
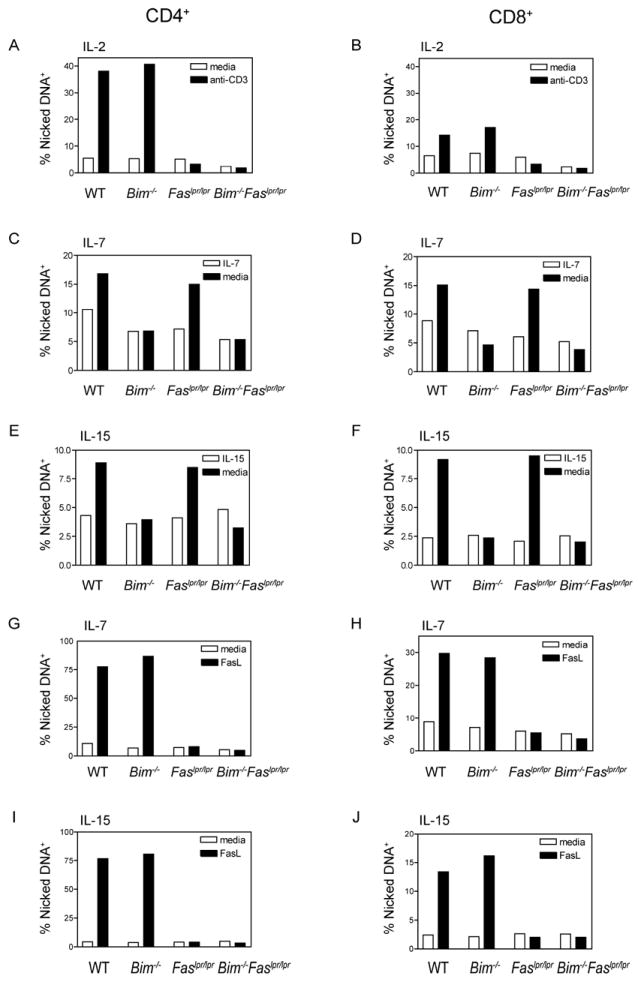
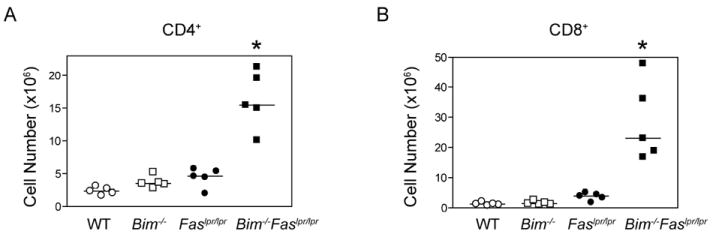
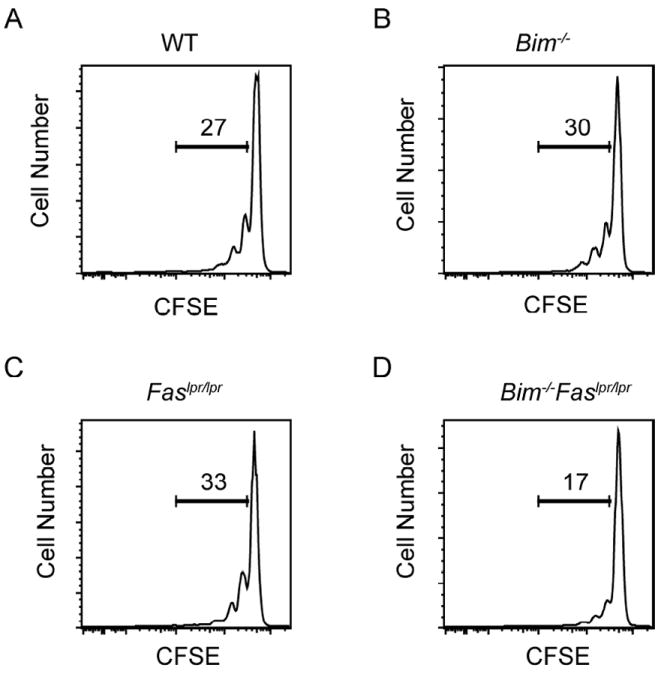
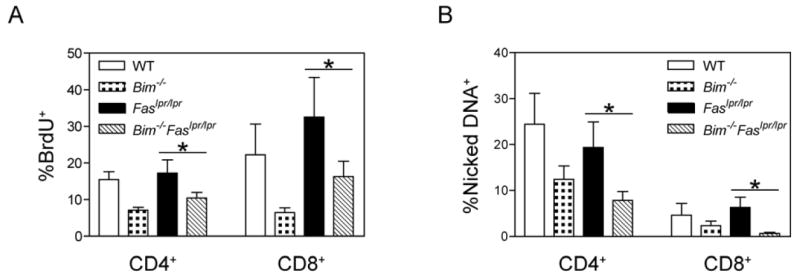
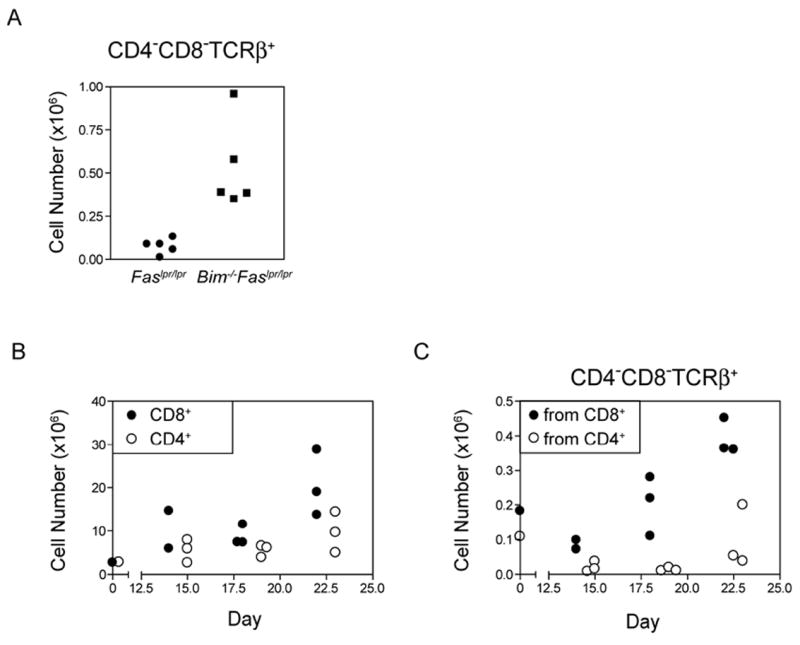
The size of the peripheral T-lymphocyte compartment is governed by complex homeostatic mechanisms that balance T-cell proliferation and death. Proliferation and survival signals are mediated in part by recurrent self-peptide/MHC-TCR interactions and signaling by the common γ chain-containing cytokine receptors, including those for IL-7 and IL-15. We have previously shown that the death receptor Fas (CD95/APO-1) regulates apoptosis in response to repeated TCR stimulation, whereas the Bcl-2 homology domain 3-only protein Bim mediates cytokine withdrawal-induced apoptosis. We therefore reasoned that these two molecules might cooperate in the regulation of homeostatic proliferation. In this study, we observe that the combined loss of Fas and Bim synergistically enhances the accumulation of T cells in lymphopenic host mice, and this is particularly pronounced for the unusual CD4(-) CD8(-) TCRαβ(+) T cells that are characteristic of Fas-deficient (Fas(lpr/lpr) ) mice. Our findings demonstrate that these CD4(-) CD8(-) TCRαβ(+) T cells arise from homeostatic proliferation of CD8(+) T cells. These studies also underscore the profound rate of baseline T-cell proliferation that likely occurs in wild-type mice even in the absence of foreign antigen, and the consequent need for its coordinated regulation by multiple death-signaling pathways.
Copyright © 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
The authors have no conflicting financial or commercial interests.
Figures
References
-
- Ernst B, Lee DS, Chang JM, Sprent J, Surh CD. The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity. 1999;11:173–181. - PubMed
-
- Ku CC, Murakami M, Sakamoto A, Kappler J, Marrack P. Control of homeostasis of CD8+ memory T cells by opposing cytokines. Science. 2000;288:675–678. - PubMed
-
- Schluns KS, Kieper WC, Jameson SC, Lefrancois L. Interleukin-7 mediates the homeostasis of naive and memory CD8 T cells in vivo. Nature Immunol. 2000;1:426–432. - PubMed
-
- Surh CD, Sprent J. Homeostasis of naive and memory T cells. Immunity. 2008;29:848–862. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
