

Jagged1 in the portal vein mesenchyme regulates intrahepatic bile duct development: insights into Alagille syndrome
- PMID: 21062863
- PMCID: PMC2976287
- DOI: 10.1242/dev.052118
Jagged1 in the portal vein mesenchyme regulates intrahepatic bile duct development: insights into Alagille syndrome
Abstract
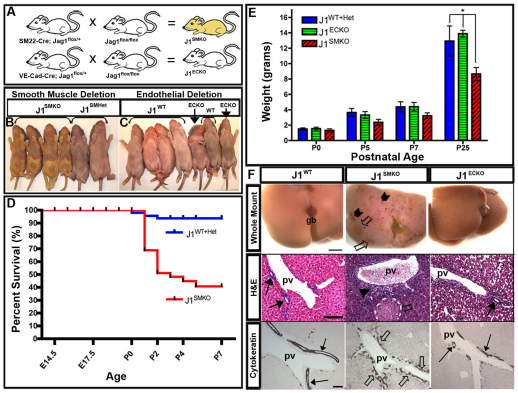
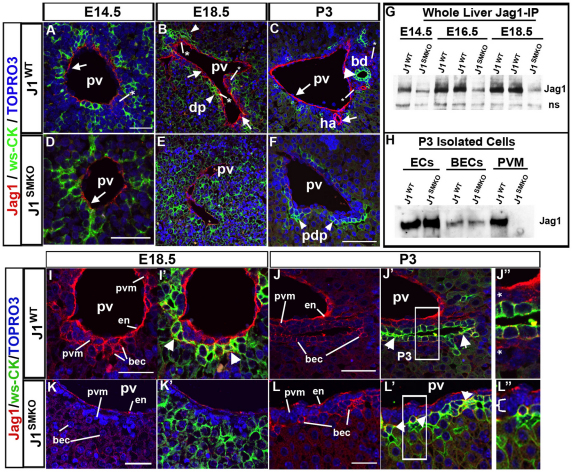
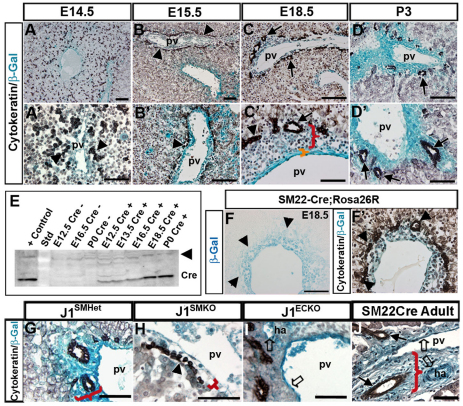
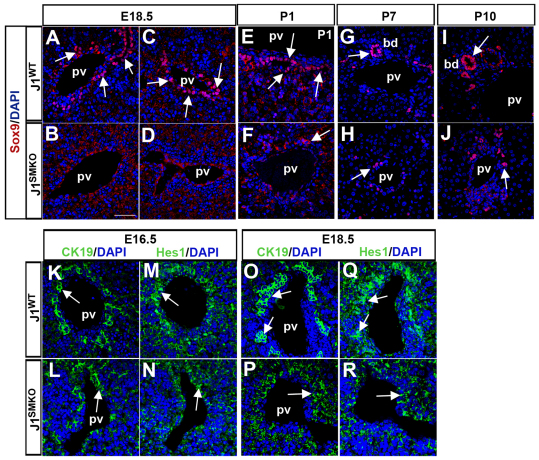
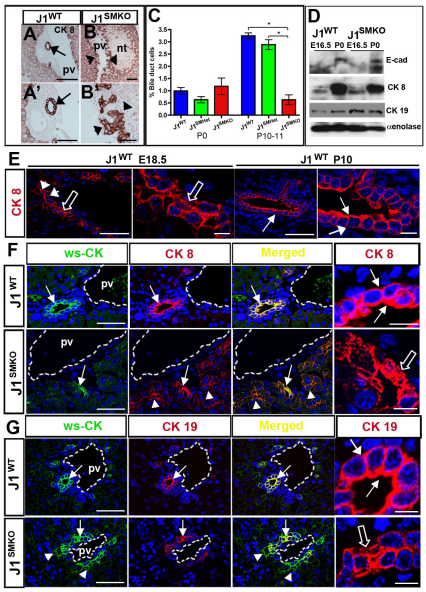
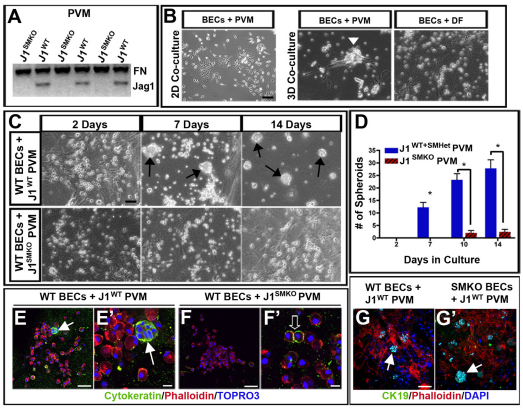
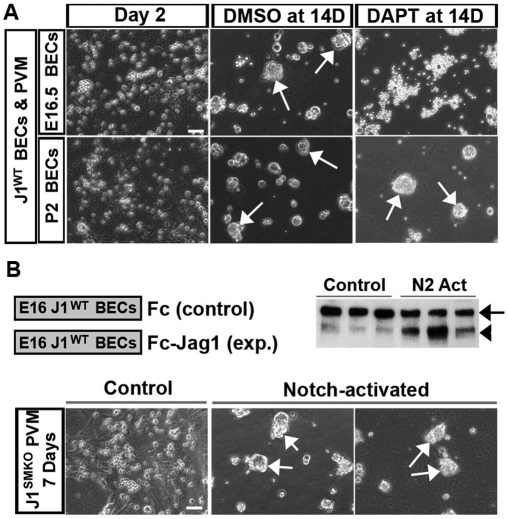
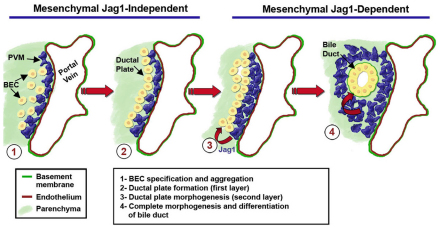
Mutations in the human Notch ligand jagged 1 (JAG1) result in a multi-system disorder called Alagille syndrome (AGS). AGS is chiefly characterized by a paucity of intrahepatic bile ducts (IHBD), but also includes cardiac, ocular, skeletal, craniofacial and renal defects. The disease penetration and severity of the affected organs can vary significantly and the molecular basis for this broad spectrum of pathology is unclear. Here, we report that Jag1 inactivation in the portal vein mesenchyme (PVM), but not in the endothelium of mice, leads to the hepatic defects associated with AGS. Loss of Jag1 expression in SM22α-positive cells of the PVM leads to defective bile duct development beyond the initial formation of the ductal plate. Cytokeratin 19-positive cells are detected surrounding the portal vein, yet they are unable to form biliary tubes, revealing an instructive role of the vasculature in liver development. These findings uncover the cellular basis for the defining feature of AGS, identify mesenchymal Jag1-dependent and -independent stages of duct development, and provide mechanistic information for the role of Jag1 in IHBD formation.
Figures
References
-
- Alagille D., Estrada A., Hadchouel M., Gautier M., Odievre M., Dommergues J. P. (1987). Syndromic paucity of interlobular bile ducts (Alagille syndrome or arteriohepatic dysplasia): review of 80 cases. J. Pediatr. 110, 195-200 - PubMed
-
- Alva J. A., Zovein A. C., Monvoisin A., Murphy T., Salazar A., Harvey N. L., Carmeliet P., Iruela-Arispe M. L. (2006). VE-Cadherin-Cre-recombinase transgenic mouse: a tool for lineage analysis and gene deletion in endothelial cells. Dev. Dyn. 235, 759-767 - PubMed
-
- Artavanis-Tsakonas S., Rand M. D., Lake R. J. (1999). Notch signaling: cell fate control and signal integration in development. Science 284, 770-776 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
