

Ethanol sensitizes mitochondria to the permeability transition by inhibiting deacetylation of cyclophilin-D mediated by sirtuin-3
- PMID: 21062897
- PMCID: PMC2987442
- DOI: 10.1242/jcs.073502
Ethanol sensitizes mitochondria to the permeability transition by inhibiting deacetylation of cyclophilin-D mediated by sirtuin-3
Retraction in
-
Retraction: Ethanol sensitizes mitochondria to the permeability transition by inhibiting deacetylation of cyclophilin-D mediated by sirtuin-3.J Cell Sci. 2016 Jul 1;129(13):2685. doi: 10.1242/jcs.193623. J Cell Sci. 2016. PMID: 27369361 Free PMC article. No abstract available.
Expression of concern in
-
Expression of Concern: Ethanol sensitizes mitochondria to the permeability transition by inhibiting deacetylation of cyclophilin-D mediated by sirtuin-3. Nataly Shulga, John G. Pastorino. J Cell Sci doi: 10.1242/jcs.073502.J Cell Sci. 2016 Feb 15;129(4):869. doi: 10.1242/jcs.187005. J Cell Sci. 2016. PMID: 26905966 Free PMC article. No abstract available.
Abstract
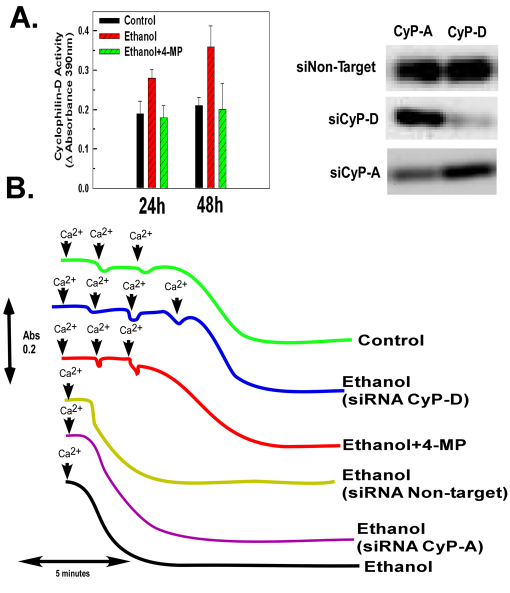
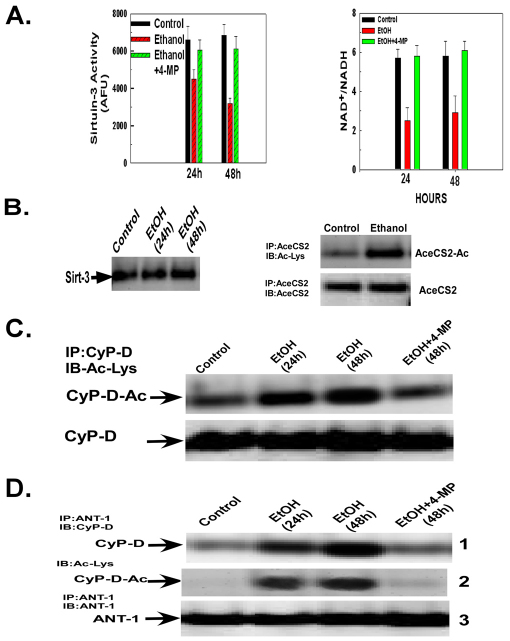
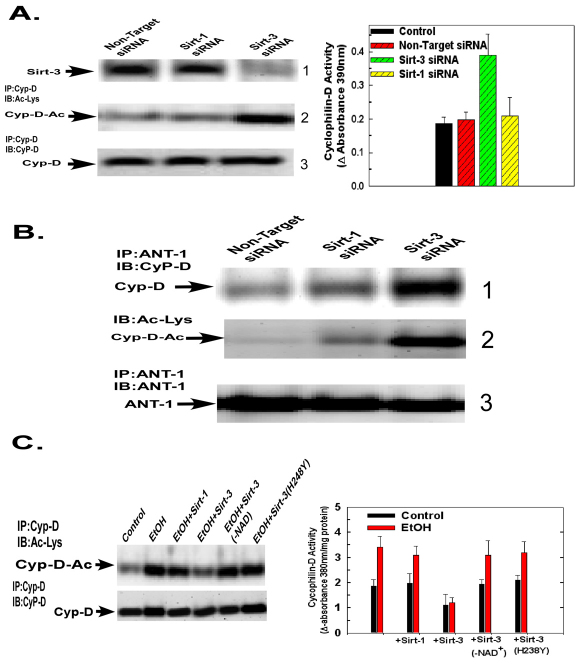
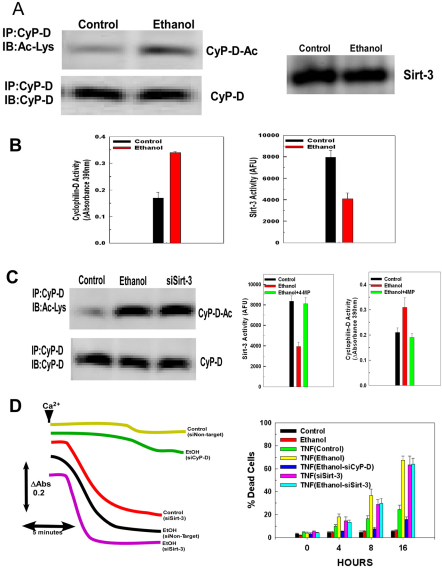
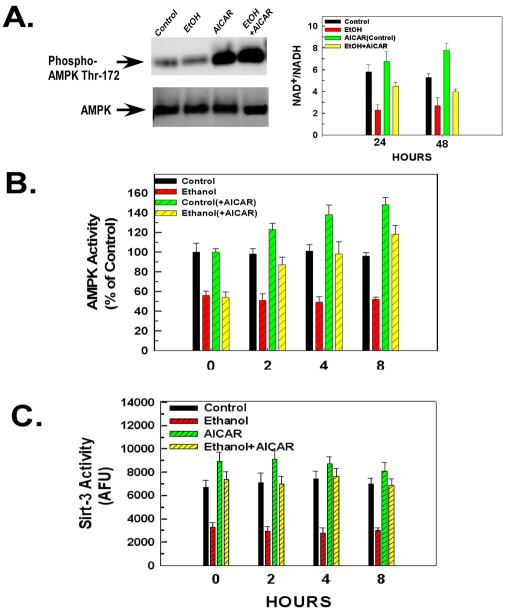
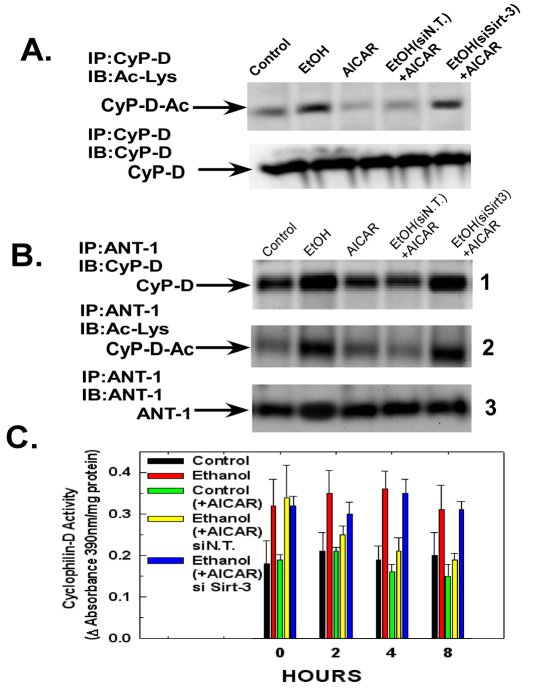
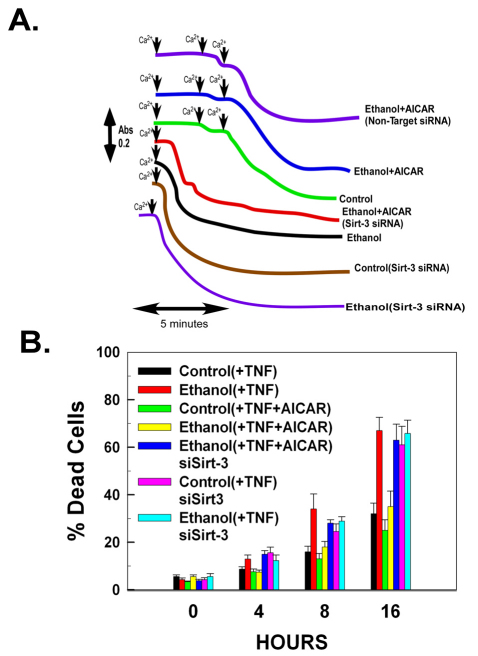
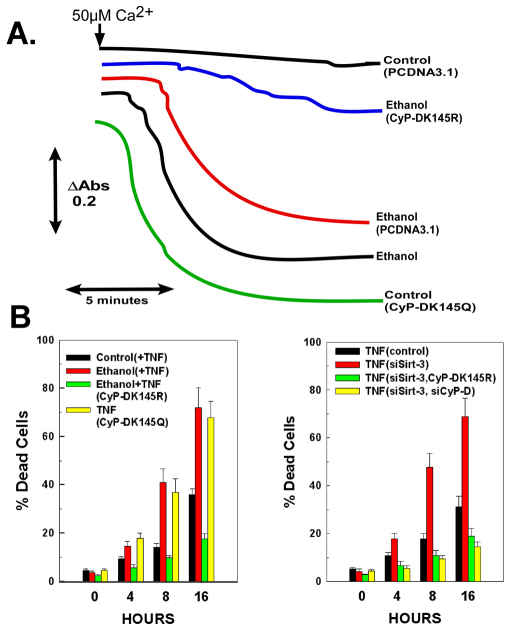
Ethanol increases the vulnerability of mitochondria to induction of the mitochondrial permeability transition (MPT). Cyclophilin-D activity enhances the potential for the permeability transition pore (PTP) to open. In the present study, we demonstrate that ethanol and its metabolism sensitize the PTP to opening, in part by increasing the acetylation and activity of cyclophilin-D. This effect of ethanol is mediated by inhibiting the activity of sirtuin-3, an NAD(+) dependent deacetylase that is localized to the mitochondrial matrix. The ethanol-enhanced acetylation of cyclophilin-D also increases the interaction of cyclophilin-D with the adenine nucleotide translocator-1 (ANT-1) and is dependent on ethanol metabolism. Moreover, activation of AMPK, a known positive modulator of sirtuin activity, prevented the ethanol-induced suppression of sirtuin-3 activity and the attendant increase of cyclophilin-D acetylation, activity and association with ANT-1. Additionally, AMPK reactivation of sirtuin-3 prevented the sensitization to the MPT and the enhancement of cell killing by TNF in cells exposed to ethanol.
Figures
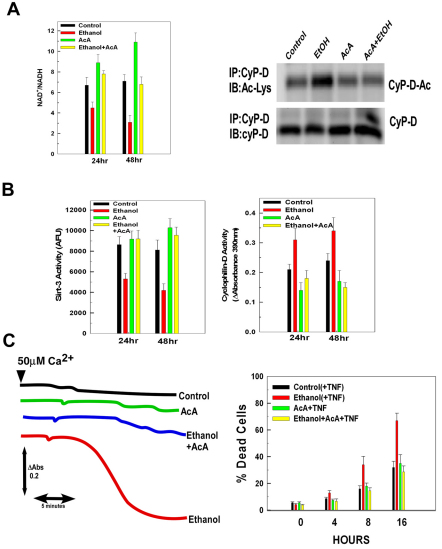
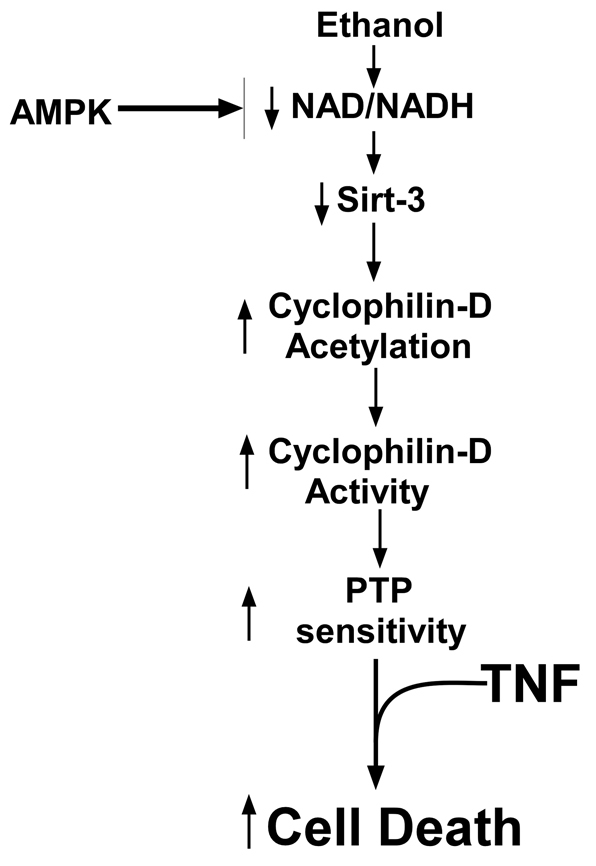
Comment in
-
Findings of research misconduct.NIH Guide Grants Contracts (Bethesda). 2016 Jun 10:NOT-OD-16-104. NIH Guide Grants Contracts (Bethesda). 2016. PMID: 27311138 Free PMC article. No abstract available.
-
Findings of Research Misconduct.Fed Regist. 2016 May 13;81(93):29871. Fed Regist. 2016. PMID: 27737277 Free PMC article. No abstract available.
References
-
- Baines C. P., Kaiser R. A., Purcell N. H., Blair N. S., Osinska H., Hambleton M. A., Brunskill E. W., Sayen M. R., Gottlieb R. A., Dorn G. W., et al. (2005). Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature 434, 658-662 - PubMed
-
- Basso E., Fante L., Fowlkes J., Petronilli V., Forte M. A., Bernardi P. (2005). Properties of the permeability transition pore in mitochondria devoid of Cyclophilin D. J. Biol. Chem. 280, 18558-18561 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
